家马起源与驯化的古DNA研究进展
发布时间:2024年11月07日
一、引言
动物的驯化作为人类发展史上最重要的转变之一[1],最早见于距今11000~4000年前狩猎采集者对狼的控制[2]。到了新石器时代至青铜时代,许多其他物种逐渐被纳入人类视野,如绵羊[3]、山羊[4]、牛[5]、猪[6]、鸡[7]和马[8]。这些动物自驯化以来,一直在人类社会扮演着多种重要角色。它们不仅能为我们提供肉蛋奶等食物资源,也被应用于祭祀、交通等方面。学者研究动物驯化的问题,主要从两个方面入手:一是从动物考古学角度,基于考古材料,利用传统手段对骨骼形态变化、死亡年龄结构、性别比例和同位素等进行分析;二是用古DNA技术手段从现代家养动物中获得遗传信息以追溯他们的驯化时间和地理起源。
马(Equus caballus)在动物分类学上属于脊椎动物亚门(Vertebrata)、哺乳纲(Mammalia)、奇蹄目(Perissodactyla)、马科(Equidae)、马属(Equus),是一种草食性动物,现存亚种有家马和普氏野马两种。在史前考古学研究中,马的驯化不仅代表了游牧文明的持续发展,也是人类文明进步的象征[9]。马在古代社会不仅可作肉食之用,更在交通运输和军事战争等方面起到重要作用,可以说马的到来促进了人群的迁徙和民族的融合。近年来,考古遗址中出土的马骨样本为我们研究家马驯化相关问题提供了时间和地点线索。本文通过对目前家马起源与驯化的考古学背景、线粒体DNA研究、Y染色体研究、全基因组研究等内容进行了系统整理,以期为未来研究提供一定的参考。
二、家马起源与驯化的考古学背景
中国家马的起源、驯化及扩散问题引起了学者的极大兴趣。自距今6000年以来,马骨遗骸已经在欧亚大陆和西伯利亚草原带有了较为广泛的分布[10]。结合我国目前的考古发现来看,从旧石器时代开始,就有马骨遗骸源源不断地出现在我们的视野当中。具体来说,旧石器时代仅存在野马,它们大多出现在我国北方地区,且常常作为被捕猎的对象[11];在甘肃、青海、陕西和河南等地的新石器时代和青铜时代早期遗址中发现了马牙和马骨碎片,但很难确定它们是否被驯化[12];到了商代晚期,黄河中下游地区出现较多的马骨遗骸,它们大多用于殉葬和祭祀[13]。但截至目前,新石器时代晚期至商代早期关于马的考古资料、文字资料仍然不甚丰富[14]。
关于中国古代家马起源的问题,目前学术界分为两种主流观点:①外来输入说,即马和马车由黑海和里海之间的中亚草原地带传入中国,经由我国西北地区进一步传至中原地区[15];②本地起源说,虽然中亚和西亚地区考古发现的家马时间上较我国早,但这并不能表明我国的家马来自于西方或北方,中国家马可能由本地的普氏野马(Equus przewalskii)作为祖先独立起源[16]。基于这两种观点,许多学者对中国古代家马的DNA进行了分析。以往的研究表明:中国家马的起源问题较为复杂,可能在经历了外来传入之后又受到本地驯化的影响,但目前尚未有证据表明中国家马和普氏野马之间存在直接的谱系关系[17]。
传统动物考古学研究家马驯化问题通常采用对马骨样本牙齿和骨骼进行形态测量的方法,进而分析死亡年龄结构、病理现象、性别和数量比例特征等[18],结合考古学文化面貌来判断出土的马骨是否属于家马范畴,再进一步研究其起源及驯化问题。但在驯化的早期阶段,野马和家马在骨骼形态上的差异并不明显,这就需要通过古DNA研究遗传多样性进而推断初始的驯化中心。家马和野马的形态差异出现在铁器时代早期(距今约3000年前),而在距今约5500年前的博泰(Botai)遗址中,已经可以找到马具、挤奶和放牧的证据[19]。最近的基因组学研究表明,博泰马是普氏野马的祖先[20]。目前学术界认为中亚草原地带可能是家马的驯化中心,这是基于当地现代家马具有较高的遗传多样性来说的,而最近的研究表明,距今4700~4000年前存在于伊比利亚的古马属于已经灭绝的马的谱系,并非现代家马的祖先[21]。由此可见,家马的起源与驯化问题仍有许多未解之谜,若想深入探讨这一问题,则需要动物考古学和古DNA研究相结合来共同完成。
三、线粒体DNA研究
线粒体DNA(mtDNA)作为古DNA研究最常用的遗传标记之一,具有多拷贝数、母系遗传、极少发生重组、进化速率快等特点,目前已被广泛应用于家马的起源、驯化及扩散的研究中。通过对家马的线粒体DNA进行研究,可以探究家马的母系来源。
(一)世界家马的线粒体DNA研究
关于世界家马的线粒体DNA研究,始于20世纪90年代,由于测序技术手段等的限制,当时的研究主要围绕线粒体DNA控制区(D-loop)片段展开[22]。2001年,卡尔斯·维拉(Carles Vilà)等通过对616个现代马的37个不同的线粒体DNA D-loop区序列进行系统发育分析,将样本划分为6个线粒体DNA谱系(A~F),发现现代家马的母系来源较为广泛,认为马的高线粒体DNA序列多样性意味着母系的广泛的、前所未有的整合以及野马的广泛利用和驯化[23]。
2002年,托马斯·詹森(Thomas Jansen)等对全球652匹马的线粒体DNA D-loop区序列进行分析构建中介网络图,将现代家马进一步划分为7个线粒体DNA谱系(A~G)。该研究也发现了几个谱系与地理分布或品种之间存在特定的对应关系:A2亚组主要由普氏野马组成;C1亚组主要存在于埃克斯穆尔高地马、挪威峡湾马、冰岛矮马、苏格兰高地马等北欧地区的矮马中;D1亚组主要存在于伊比利亚和北非,如安达卢西亚马、卢西塔诺马和柏布马等[24]。随后,安吉拉·麦加亨(Angela McGahern)等对118匹来自东方马群体的线粒体DNA进行了AMOVA方差分析,发现单倍群F是欧亚大陆东部的家马群体的主要组成部分,再一次印证了在马线粒体DNA序列中可以检测到较为明显的地理分布规律[25]。
2010年,迈克尔·切斯拉克(Michael Cieslak)等为了进一步解决家马的线粒体DNA高度变异以及母系遗传的起源和历史等问题,对来自207个古代遗址和1754个现代马的1961个线粒体DNA序列进行分析,这些样本的时空分布非常广泛,从阿拉斯加、北西伯利亚和东西伯利亚到伊比利亚半岛,时间跨度从晚更新世到现代。该研究根据线粒体DNA控制区数据显示在铁器时代有多个母系的驯化和基因的深入,结合系统发育网络分析将家马更详细地划分为A~K和X1~X7的19个单倍群,进一步揭示了古代马线粒体谱系的地理起源[26]。
随着测序手段的发展,线粒体DNA研究从控制区片段发展到线粒体全基因组研究,这使得我们能够获得更多有效的遗传信息。卢多维奇·奥兰多(Ludovic Orlando)等人在2011年利用Helicos HeliScope和Illumina GAIIx平台对一块更新世的马骨遗骸进行了单分子测序,该研究利用的二代测序技术(NGS)对古代家马基因组的研究具有里程碑式的意义[27]。随后,塞巴斯帝安·利波德(Sebastian Lippold)等对44个品种的59匹家马和1匹普氏野马的线粒体基因组通过微阵列捕获与高通量测序技术进行分析,认为现代马在距今9.3万年前和距今3.8万年前存在共同的祖先,贝叶斯天际线图(BSP)揭示了从距今8000到6000年前开始了显著的种群扩张,大量野马的线粒体基因组被纳入了家马种群,证实了现代家马的母系来源多样性[28]。
2012年,亚历山德罗·阿奇利(Alessandro Achilli)等分析了来自亚洲、欧洲、美洲和中东的83匹现代马的线粒体全基因组数据,将马线粒体DNA系统发育提高到分子分辨率的最高水平。该研究划分了18个主要的单倍群(A~R),其中在亚洲的现代马中检测到了所有的单倍型,且单倍群F仅在普氏野马中发现,表明在全球范围内的现代家马具有非常丰富的线粒体遗传多样性(图一)[29]。
通过对世界范围内家马线粒体DNA的研究,现有的成果大多支持家马的母系来源十分广泛这一观点。在研究家马驯化相关问题的过程中,随着对马骨样本形态学研究和古DNA研究的深入,也有新的马亚属成员逐渐被发现,其中较为重要的就是Equus(Sussemionus)[31]。结合化石证据来看,Equus(Sussemionus)这一亚属进化得相当成功,对于干旱或潮湿的环境均能适应,且其分布范围非常广泛,在美洲、非洲和西南西伯利亚均有发现[32]。这一世系最初被学者认为在中更新世已经灭绝,因为当时发现的最后一枚典型的Equus(Sussemionus)样本的年代为距今50万年前[33]。然而在最近的研究中,有学者发现了类似形态化石的存在。2011年,维拉·艾森曼(Véra Eisenmann)等在俄罗斯西伯利亚西南部哈卡西亚共和国(Khakassia)晚更新世的普罗斯库里亚科娃(Proskuriakova)洞穴中发现了一种新的未驯化的马,其年代可以追溯到距今4万年前。经过古DNA研究,确定了它是属于Equus(Sussemionus)亚属中的一员——Equus(Sussemionus)ovodovi(奥氏马)[34]。2017年,安娜·S.德鲁日科娃(Anna S.Druzhkova)等分析了来自普罗斯库里亚科娃洞穴的马骨样本和阿尔泰地区丹尼索瓦(Denisova)洞穴的马骨样本的线粒体全基因组数据,发现奥氏马是与家马、驴马、斑马并列的亚属,并将其存在的时间下限延伸到距今3.2万年左右[35]。
(二)中国家马的线粒体DNA研究
关于中国家马的线粒体DNA研究,同样始于20世纪90年代,当时的研究主要集中在酶切多态性研究。到了21世纪,随着一代测序技术的发展,学者逐渐将目光转向线粒体DNA D-loop区的研究。芒来等在2005年通过对比现生的中国蒙古马和国外纯血马的线粒体DNA D-loop区序列,发现这两个品种的马匹具有一定的相似性,且线粒体多态性均很丰富[36]。
2007年,蔡大伟等对来自西藏拉萨和泽当的23匹西藏现代马线粒体DNA D-loop区序列进行分析,认为西藏马的母系遗传多样性较为丰富,且西藏马在母系遗传方面与近东、中亚以及欧洲家马有相近的亲缘关系而与韩国马亲缘关系较远[37];对来自内蒙古赤峰地区大山前遗址和井沟子遗址的9个青铜时代马骨遗骸进行了线粒体DNA 16S rRNA和D-Loop区序列分析,通过构建系统发育网络图,发现赤峰地区的古马具有较高的母系遗传多样性,同时也反映出中国家马起源问题的复杂性[38]。
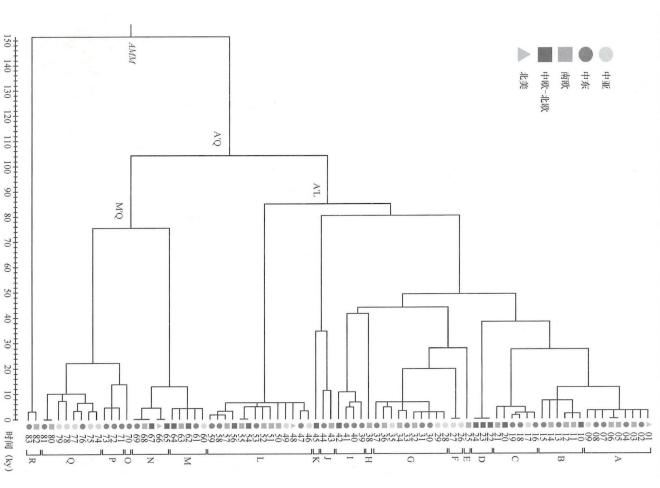
图一 基于线粒体全基因组的现代马系统发育树[30]
2008年,蔡大伟等对内蒙古凉城县小双古城墓地和板城墓地的11个春秋时期马骨样本的线粒体DNA D-Loop区序列进行分析,研究表明中国家马的母系来源非常广泛,认为在家马驯化的过程中,有来自其他地区马的基因流的引入[39]。同年,孙锐等对陕西关中的27匹马进行了线粒体DNA D-loop区遗传多态性和系统进化的分析,发现关中马存在A、C、D、F和G共5个谱系,表明关中马的起源同样具有多样性[40]。
2009年,蔡大伟等对中国北方9个考古遗址出土的35个马骨遗骸进行线粒体DNA研究,得出中国古代家马母系遗传十分多样的结论,且中国古代马与现代蒙古马的关系更为密切。研究指出单倍型F可能是一个古老的东亚起源单倍群,表明中国家马起源的复杂性,并且在初始驯化后可能有外部线粒体DNA输入(图二)[41]。同年,雷初朝等分析了来自中国各地的182匹现代马的线粒体D-loop区序列,发现世系A~G在中国现代家马中均有分布,暗示中国家马的起源驯化过程具有复杂性,支持中国家马的多重起源[42]。
2012年,张涛等对中国西部3个本土马品种(宁强马、贵州马和哈萨克马)43匹马的线粒体DNA D-loop区序列构建系统发育树,证实这三个本土马品种具有多个母系来源[44]。
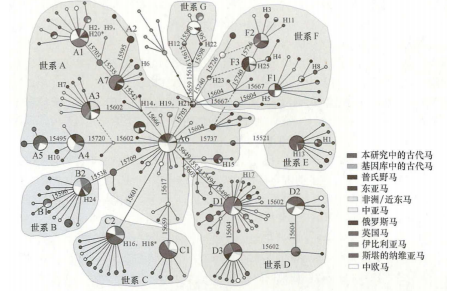
图二 家马线粒体单倍型中介网络图[43]
2014年,赵欣等对新疆石人子沟西汉墓葬出土的马骨进行古DNA分析,认为该遗址出土的家马具有高度线粒体遗传多样性,推测新疆因其特殊的地理位置,在外域家马传入中国过程中起到非常重要的作用[45];2015年该课题组又对陕西淳化枣树沟脑遗址西周中晚期马坑出土的马骨遗骸进行线粒体DNA D-loop区研究,认为该遗址的马同样具有较高的线粒体DNA遗传多样性,且与中国北方地区尤其是内蒙古长城地带的家马母系亲缘关系较近[46]。随后,赵欣等又对来自新疆木垒县平顶山墓群的8个青铜时代中晚期的马骨样本进行线粒体DNA D-loop区分析,发现这些样本同样具有较高的母系遗传多样性,且均为家马[47]。
2017年,杨云周等分析了来自中国25个地方品种共计714匹马的线粒体DNA D-loop区序列,发现了新的变异并将之命名为H和I世系,遗传多样性结果表明引进的品种可能从欧亚草原通过北方地区进入中国[48]。
2018年,蔡大伟等通过对陕西凤翔秦公一号大墓车马坑出土的10个古代马进行古DNA分析,认为部分古代马的线粒体DNA谱系在欧亚大陆西部起源;同时发现该遗址个别古代马(FX4和FX5)的基因型与汗血马有关,推测汗血马可能早在西周时期就已经通过贸易手段来到我国西北地区[49]。
2019年,赵欣等分析了青海省民和县喇家遗址出土的8个汉代马骨样本的线粒体DNA数据,发现它们可以归属到3个不同的谱系(A、D和F),表明西北甘青地区是外域家马进入中国的一条主要路线[50]。同年,蔡大伟等对宁夏隆德县沙塘北塬遗址出土的5个新石器时代马骨样本进行线粒体DNA分析,结果发现了来自西方的C1单倍型[51];朱司祺等对新疆塔城松树沟遗址的2匹早期铁器时代的古代马进行线粒体DNA分析,推测该遗址的古马从欧亚草原经由阿尔泰地区传入国内,进一步表明我国古代家马的驯化与西方因素的传入密切相关[52]。
2020年,文少卿等对陕西凤翔雍山血池秦汉皇家祭祀遗址出土的26个马骨样本进行线粒体DNA研究,结果显示它们可以归属到6个线粒体谱系(A~F)的11个单倍群,说明其具有很高的母系来源多样性,且与现代蒙古马更为接近[53]。同年,东晓玲等对甘肃省礼县西山遗址的3匹家马骨骼进行了线粒体DNA分析,发现该遗址马骨同样具有母系遗传多样性,为研究家马驯化早期引入中国的路线提供了新的数据[54]。
2022年,李鹭分析了内蒙古自治区包头燕家梁遗址13个元代马骨样本的线粒体全基因组数据,证实了该遗址的古马具有丰富的母系遗传多样性,推测燕家梁遗址作为驿站在中国古代家马的驯化传播中起到连接作用[55]。同年,张新雨提取了陕西靖边庙梁遗址出土的17个新石器时代马骨样本的线粒体全基因组数据,结合中国北方三个遗址(洪河遗址、木柱柱梁遗址、沙塘北塬遗址)中距今4000年左右马骨样本的线粒体全基因组数据进行分析,发现该遗址的样本属于奥氏马。研究表明在奥氏马群体内部可能存在两个分支:EOV1分支以陕西地区的样本为主,EOV2以黑龙江及西伯利亚地区的样本为主,但由于缺乏内蒙古地区的奥氏马样本,奥氏马的扩散传播路线尚不清楚[56]。
四、Y染色体研究
由于线粒体DNA仅能反映出母系方面的遗传信息,不能解释父系遗传的相关信息,若想研究家马的父系起源问题,则需要对马的Y染色体进行分析。
2003年,芭芭拉·沃尔纳(Barbara Wallner)等首次对家马的Y染色体进行分析,研究发现普氏野马和家马属于姊妹分类群,普氏野马并非家马的父系祖先[57]。随后,该课题组使用代表性差异分析(RDA)成功获得6个家马Y染色体微卫星标记(SSR),发现Y染色体微卫星标记在家马中显示出单一的单倍型[58]。
2004年,加布里埃尔·林格伦(Gabriella Lindgren)等分析了来自15个品种的52匹雄性马匹的Y染色体序列,发现与家马广泛的母系来源多样性相比,家马的父系祖先十分有限,可能是在驯养过程中加入了性别选择[59]。
2010年,凌英会等对来自中国的家马(30个品种)、普氏野马和驴的573个雄性样本的父系遗传变异进行了6个Y染色体特异性微卫星测试,发现马和驴之间的Y染色体标记存在显著差异。该研究是对马Y染色体变异的首次描述,在中国家马品种中发现的2个Y染色体单倍型有助于揭开其错综复杂的遗传谱系[60]。
2012年,毛春春对来自中国的13个家马品种和国外23个家马品种共计905个家马个体进行Y染色体分析,发现中国家马有第二个父系起源的存在,进一步证实了中国家马是多父系起源的这一观点[61]。
2013年,芭芭拉·沃尔纳等使用高通量测序技术对来自欧洲的516匹雄性马匹的Y染色体的SNP分子标记进行分析,结果显示现代马Y染色体的核苷酸变异性极低,绝大部分现代马都来自6个单倍型(HT1~HT6),其中HT1是最广泛的祖先型,其他5个单倍型是由HT1诱变或驯化后的基因转化产生的。研究发现HT2和HT3在现代欧洲马种中广泛分布,HT3与英国纯种马有很高的关联性。该研究作为家马线粒体遗传谱系的补充,揭示了现代家马的父系遗传谱系[62]。
2015年,韩浩园等对来自中国西北和西南地区13个品种的304匹雄性马匹进行分析,结果显示中国马的Y染色体多态性模式与欧洲和美国马不同,但中国家马Y染色体遗传同样具有多样性,来自5个不同的Y-SNP单倍型[63]。
2017年,芭芭拉·沃尔纳等通过筛选来自21个品种的52匹马的1.46Mb Y染色体(MSY)的特异性区域,发现现代马种群的Y染色体由来自近东地区的驯化后的单一祖先产生,表明在性别导向选择中增加了来源于东方的Y染色体谱系,且英国纯种马的创始种马可以追溯到土库曼斯坦的起源[64]。
2022年,文少卿等对新疆阿勒泰地区依希根墓地出土的隋唐时期马骨样本进行了Y染色体分析,发现该遗址的样本属于由rAX突变定义的一个新分支,在父系遗传方面不属于现有的欧亚大陆西部的家马谱系,为研究中国古代家马的父系起源提供了新的例证[65]。
五、全基因组研究
在过去的10年里,二代测序技术的发展为我们带来了技术的革新,也为研究家马起源及驯化问题提供了新的思路,在对早期驯化阶段进行更深入研究的同时也着眼于思考现代育种实践对家马群体的影响。
2013年,卢多维奇·奥兰多(Ludovic Orlando)等展示了一个1.12倍覆盖率的古马基因组草图,该样本的年代可以追溯到距今约560万~78万年前。结果显示当代马、斑马和驴的分歧时间在距今450万~400万年前,是传统上认为的马属最近共同祖先分化时间的两倍。研究表明普氏野马和家马种群在距今7.2万~3.8万年前存在差异,且并未在二者之间发现基因混合的证据[66]。
2014年,迈克尔·舒伯特(Mikkel Schubert)等测得了来自俄罗斯泰米尔(Taymyr)的两个完整的古代马基因组数据,将之与家马和普氏野马的基因组进行了比较,发现晚更新世欧亚大陆的遗传结构、古代种群对现代家马的遗传变异做出了重大贡献[67]。
2015年,巴布罗·利布拉多(Pablo Librado)等分析了来自晚更新世时期的2匹雅库特古代马和9匹现代马的全基因组数据,发现现代与古代雅库特马之间不存在基因联系,可能发生过种群替代[68]。2017年,巴布罗·利布拉多等又分析了14匹来自青铜时代和铁器时代古代马的基因组数据,发现了支持神经嵴假说的早期驯化选择模式。这种模式为常见的家养特征提供了统一的发育起源,据此推测现代家马从数量有限的种马中繁殖而来[69]。
2018年,查琳·高尼茨(Charleen Gaunitz)等获得了42个古马基因组数据,其中20个来自博泰马。通过与46个已发表的古代和现代马基因组数据进行对比,发现普氏野马是在博泰放牧的马的野生后代,而非真正的野马。在距今约4000年前到现代所有家马基因组数据中,仅有约2.7%的博泰相关血统,表明大规模的基因组更替支撑了马种群的扩张,进而导致现代家马种群的出现,这一结果与青铜时代早期人口的大规模扩张相吻合[70]。
2019年,安东尼·法赫斯(Antoine Fages)等对278个古代马的基因组数据进行研究,发现在距今约5000年前的驯化早期阶段存在伊比利亚(欧亚大陆最西部)和西伯利亚(欧亚大陆最东部)已灭绝的家马世系,但这两个世系对现代家马的基因流没有贡献。在驯化后期阶段,萨珊王朝的扩张导致波斯马向欧洲和中亚的扩散,改变了3~7世纪马的种群结构和基因组成,与此同时,现代家马的遗传多样性随着现代育种实践有显著下降的趋势[71]。
2021年,巴布罗·利布拉多等收集了所有疑似驯化中心的马骨遗骸,包括伊比利亚、安纳托利亚以及欧亚大陆西部和中亚草原地带。根据273匹古代马的基因组数据,研究者认为欧亚大陆西部伏尔加河下游地区是现代家马的故乡。通过对种群变化的研究,发现现代家马的祖先群体在距今4200~4000年首先到达安纳托利亚、多瑙河下游、波希米亚和中亚,随后很快到达西欧和蒙古,最终在距今3500~3000年取代了所有当地古马群体。该研究结果否定了以前认为的博泰马是现代家马祖先的这一结论[72]。
2022年,蔡大伟等对全新世中国北方的26个马骨样本进行测序,发现它们均为奥氏马。该研究提出了奥氏马的第一个高质量的全基因组数据,测序覆盖深度高达13.4倍。系统发育分析表明,奥氏马在距今270万~230万年前从非马亚属的祖先中分化出来,并存活至全新世晚期(图三)[73]。
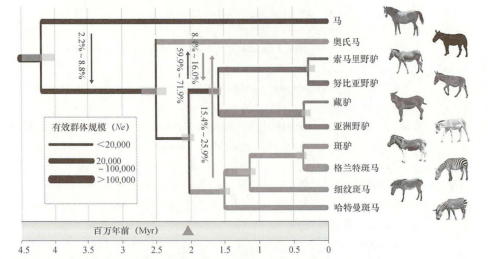
图三 已灭绝和现存马科动物的系统发育树[74]
六、结语
总结来说,目前关于家马起源与驯化问题的研究,多数认为家马的母系来源十分广泛,在驯化的过程中有野马种群的基因流不断渗入,家马的起源地点为欧亚大陆西部伏尔加河下游地区;相较于家马广泛的母系来源,多数学者的研究证实了家马父系来源较为单一,推测这属于一种管理策略——选择更为温顺可控的种马来进行繁殖,这体现了人们在驯化马的过程中加入了性别的选择。
关于我国家马驯化及扩散的问题,大多学者的研究支持中国家马由西方引入同时受到本地驯化因素的影响这一观点,并且存在两条可能的传播路线:①从欧亚大陆西部经由我国西北地区传入国内,②从欧亚大陆西部经由我国内蒙古地区传入国内。
在研究家马起源与驯化问题的过程中也有新的马亚属动物逐渐被发现,如近几年发现了存活到全新世晚期的奥氏马,对于了解中国野马的种类构成、分布范围和环境的适应性十分重要。
在过去的十年里,家马的全基因组研究极大地改变了我们对家马驯化过程的理解。动物考古学手段与古DNA研究的结合在未来能更准确地解释家马驯化的动态历史,同时对现代复杂社会产生一些新的认识。目前仍有赖于新材料的发现和新技术的突破,以便更深入地探索中国古代家马的起源和扩散问题。关于这一问题,包括传入途径在内的未解课题还有很多,还需要从多方面多角度进行深入探索,定能为家马起源与驯化研究打开新的篇章。
[1] a.Zeder M A.The domestication of animals [J].Journal of Anthropological Research, 2012, 68(2):161-190.b.Vigne J D.The origins of animal domestication and husbandry:a major change in the history of humanity and the biosphere [J].Comptes Rendus Biologies, 2011, 334(3):171-181.[2] Larson G,Karlsson E K, Perri A, et al.Rethinking dog domestication by integrating genetics archeology, and biogeography [J].Proceedings of theNational Academy of Sciences of the United States of America, 2012, 109(23):8878-8883.[3] Naderi S, Rezaei HR, Pompanon F, et al. The goat domestication process inferred from large-scale mitochondrial DNA analysis of wild and domestic individuals [J].Proceedings of the National Academy of Sciences of the United States of America,2008,105(46):17659-17664.[4] Luikart G,Gielly L,Excoffier L,et al.Multiple maternal origins and weak phylogeographic structure in domestic goats [J].Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(10):5927-5932.[5] Hongo H, Pearson J, Oksüz B, et al. The process of ungulate domestication at Çayönü, Southeastern Turkey: a multidisciplinary approach focusing on Bos sp. and Cervus elaphus [J]. Anthropozoologica, 2009, 44(1):63-78.[6] Larson G, Dobney K, Albarella U, et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication [J]. Science, 2005, 307 (5715):1618-1621.[7] Frantz L, Meijaard E, Gongora J, et al. The evolution of Suidae[J].Annual Review of Animal Biosciences, 2016, 4:61-85.[8] Zeder, MA.The origins and spread of domestic animals in Southwest Asia and Europe [J] Journal of Anthropological Research, 2015, 71(1):104-105.[9] Vilà C, Leonard J A, Gotherstrom A, et al. Widespread origins of domestic horse lineages [J]. Science, 2001, 291(5503):474-477.[10] Frantz LAF,Bradley DG, Larson G, et al.Animal domestication in the era of ancient genomics [J]. Nature Reviews Genetics, 2020, 21(8):449-460.[11] Mashkour M. Equids in time and space [C]. Papers in Honour of Vera Eisenmann(Proceedings of the 9th ICAZ Conference, Durham, 2002). Oxford: Oxbow Books, 2006(240):92-147.[12] 陈星灿.也谈家马的起源及其他[J].中国文物报,1999-6-23(3).[13] 袁靖.中国古代家马的研究[C].中国史前考古学研究祝贺石兴邦先生考古半世纪暨八秩华诞文集.西安:三秦出版社,2004:451-458.[14] 蔡大伟.古DNA与中国家马起源研究[M].北京:科学出版社,2021:25.[15] a.袁靖.中国古代家养动物的动物考古学研究[J].第四纪研究,2010(2):298-306.b.王宜涛.也谈中国马类动物历史及相关问题[J].中国文物报,1998-8-12(3).[16] a.王志俊,宋澎.中国北方家马起源问题的探讨[J].考古与文物,2001(2):26-30.b.〔美〕斯坦利.J·奥尔森著,殷志强译.中国北方的早期驯养马[J].考古与文物,1986(01):89-91.[17] Yuan J. A Zooarchaeological study on the origins of animal domestication in ancient China [J].Chinese Annals of History of Science and Technology, 2021, 5(1):1-26.[18] 袁靖.中国动物考古学[J]. 文物,2015(9):92.[19] Outram AK, Stear NA, Bendrey R, et al. The earliest horse harnessing and milking [J]. Science 2009, 323(5919): 1332-1335.[20] Gaunitz C, Fages A, Hanghaj K, et al. Ancient genomes revisit the ancestry of domestic and Przewalski's horses [J]. Science, 2018, 360(6384): 111-114.[21] Librado P, Khan N, Fages A, et al. The origins and spread of domestic horses from the Western Eurasian steppes [J]. Nature, 2021, 598(7882):634-640.[22] Xu X, Arnason U. The complete mitochondrial DNA sequence of the horse, Equus caballus: extensive heteroplasmy of the control region[J].Gene, 1994, 148(2):357-362.[24] Jansen T, Forster P, Levine MA, et al. Mitochondrial DNA and the origins of the domestic horse[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(16): 10905-10910.[25] McGahern A, Bower M A M, Edwards C J, et al. Evidence for biogeographic patterning of mitochondrial DNA sequences in Eastern horse populations [J]. Anim Genet, 2006, 37(5):494- 497.[26] Cieslak M, Pruvost M, Benecke N, et al. Origin and history of mitochondrial DNA lineages in domestic horses [J]. PLoS One, 2010, 5(12): e15311.[27] Orlando L, GinolhacA, RaghavanM, et al. True single-molecule DNA sequencing of a pleistocene horse bone [J]. Genome Research, 2011, 21(10): 1705-1719.[28] Lippold S, MatzkeN J, Reissmann M, et al. Whole mitochondrial genome sequencing of domestic horses reveals incorporation of extensive wild horse diversity during domestication [J].BMC Evolutionary Biology, 2011, 11(1):328.[29] Achilli A, Olivieri A, Soares P, et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication [J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(7):2449-2454.[31] Eisenmann V. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia)[J]. Comptes Rendus Biologies, 2010, 333(3): 235-240.[32] a.朱司祺.中国北方三个遗址出土马属动物(Equus ovodovi)的分子考古学研究[D]. 吉林大学博士学位论文,2020.b.同[31] .[33] Vasiliev S K. Large Mammal Fauna from the Pleistocene deposits of Chagyrskaya Cave Northwestern Altai(based on 2007-2011 excavations)[J]. Archaeology, Ethnology and Anthropology of Eurasia, 2013,41(1):28-44.[34] a.Eisenmann V, Sergej V. Unexpected finding of a new Equus species(Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in late Pleistocene Deposits of Khakassia(Southwestem Siberia)[J]. Geodiversitas, 2011, 33:519-530.b.Orlando L, Metcalf J L, Alberdi M T, et al.Revising the recent evolutionary history of equids using ancient DNA[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(51):21754-9.c.Vilstrup JT, Seguin-Orlando A, Stiller M, et al. Mitochondrial phylogenomics of modern and ancient equids [J]. PLoS One, 2013, 8(2):e55950.[35] Druzhkova AS, AI Makunin, NV Vorobieva, et al. Complete mitochondrial genome of an extinct Equus(Sussemionus)ovodovi specimen from Denisova cave(Altai, Russia) [J]. Mitochondrial DNA PartB, 2017, 2(1):79-81.[36]芒来,李金莲,石有斐.中国蒙古马与国外纯血马mtDNA D-loop高变区序列比较[J].遗传,2005(1):91-94.[37]蔡大伟,韩璐,李胜男等.西藏马线粒体DNA D-loop区的遗传多样性[J].吉林大学学报(理学版),2007,45(5):873-878.[38]蔡大伟,韩璐,谢承志等.内蒙古赤峰地区青铜时代古马线粒体DNA分析[J].自然科学进展,2007(3):385-390.[39]蔡大伟,曹建恩,陈全家等.内蒙古凉城县春秋时期古代马线粒体DNA分析[C].边疆考古研究(第7辑).北京:科学出版社,2008:328-333,447.[40]孙锐,谢文美,张云生等.关中马mtDNA D-loop区遗传多态性分析[J].西北农业学报,2008(6):17-20.[41] Cai D, Tang Z, Han L, et al. Ancient DNA provides new insights into the origin of the Chinese domestic horse [J]. Journal of Archaeological Science, 2008, 36(3):835-842.[42] Lei CZ, Su R, Bower M A, et al. Multiple maternal origins of native modern and ancient horse populations in China[J]. Animal Genetics, 2009, 40(6):933-944.[44] Zhang T, Lu H, Chen C, et al.Genetic Diversity of mt DNA D-loop and Maternal Origin of Three Chinese Native Horse Breeds [J]. Asian-Australasian Journal of Animal Sciences, 2012, 25(7):921-926.[45] 赵欣,Antonia T R,尤悦等.新疆石人子沟遗址出土家马的DNA研究[J].第四纪研究,2014(1):187-195.[46] 赵欣,李悦,陈洪海等.陕西省淳化县枣树沟脑遗址马坑出土马骨的DNA初步研究[J].南方文物,2015(3):70-76.[47] 赵欣,东晓玲,韩雨等.新疆木垒县平顶山墓群出土马骨的DNA研究[J].南方文物,2017(3):187-191.[48] Yang Y, ZhuQ, LiuS, etal. The origin of Chinese domestic horses revealed with novel mtDNA variants[J]. Anim Sci J, 2017, 88(1): 19-26.[49] 蔡大伟,朱司祺,胡松梅等.陕西凤翔秦公一号大墓车马坑马骨遗骸古DNA研究[J].考古与文物,2018(3):106-112.[50] 赵欣,吕鹏,东晓玲等.青海省民和县喇家遗址出土汉代马骨的DNA初步研究[C].南方文物,2019(4):187-195.[51] 蔡大伟,李畅,朱司祺等.宁夏隆德县沙塘北塬遗址出土马骨的DNA分析[C].边疆考古研究(第26辑).北京:科学出版社,2019:303-310.[52] 朱司祺,蔡大伟,张乃凡等.新疆塔城松树沟遗址出土古代马的线粒体基因组分析[C].边疆考古研究(第26辑).北京:科学出版社,2019:311-320.[53] 文少卿,俞雪儿,孙畅等.凤翔雍山血池遗址北斗坊7号坑马骨的古DNA研究[J].考古与文物,2020(6):115-121.[54] 东晓玲,赵欣,吕鹏等.甘肃省礼县西山遗址出土马骨的DNA初步研究[J].南方文物,2020(4):182-186.
[55] 李鹭.包头燕家梁遗址出土古代马属动物的线粒体全基因组分析[D].吉林大学硕士学位论文,2022.[56] 张新雨.陕西靖边庙梁遗址出土古代马属动物的分子考古学研究[D].吉林大学硕士学位论文,2022.[57] Wallner B, Brem G, Muller M, et al. Fixed nucleotide differences on the Y chromosome indicate clear divergence between Equus przewalskii and Equus caballus [J]. Anim Genetics, 2003, 34(6): 453-456.[58] Wallner B, Piumi F, Brem G, et al. Isolation of Y chromosome-specific microsatellites in the horse and cross-species amplification in the genus Equus [J]. Journal of Heredity, 2004, 95(2): 158- 164.[59] Lindgren G, Backstrom N, Swinburme J, et al.Limited number of patrilines in horse domestication [J].Nature Genetics, 2004, 36(4): 335-336.[60] Ling Y, Ma Y, Guan W, et al. Identification of Y chromosome genetic variations in Chinese indigenous horse breeds [J]. Journal of Herediy, 2010, 101(5): 639-643.[61] 毛春春.马KIT基因与Y染色体遗传多样性研究[D]. 西北农林科技大学硕士学位论文,2012.[62] Wallner B, Vogl C, Shukla P, et al.Identification of genetic variation on the horse y chromosome and the tracing of male founder lineages in modern breeds [J]. PLoS One, 2013, 8(4): e60015.[63] Han H, Zhang Q, GaoK, et al. Y-Single nucleotide polymorphisms diversity in Chinese indigenous horse [J]. Asian-Australasian Jourmal of Animal Sciences, 2015, 28(8):1066-1074.[64] Wallner B, Palmieri N, Vogl C, et al. Y Chromosome uncovers the recent oriental origin of modern stallions [J].Current Biology, 2017, 27(13):2029-2035.[65] 文少卿,于建军,魏偏偏等.新疆阿勒泰地区依希根墓地出土马骨的分子考古研究[J]. 西域研究,2022(1):55-65+170-171.[66] Orlando L, Ginolhac A, Zhang GJ, et al. Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse [J].Nature, 2013, 499(7456): 74-78.[67] Schubert M, Jónsson H, Chang D, et al. Prehistoric genomes reveal the genetic foundation and cost of horse domestication[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(52): E5661-5669.[68] Librado P, DerSarkissian C, ErminiL, et al. Tracking the origins of Yakutian horses and the genetic basis for their fast adaptation to subarctic environments [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(50): E6889-E6897[69]Librado P, Gamba C, Gaunitz C, et al. Ancient genomic changes associated with domestication of the horse [J]. Science, 2017, 356(6336):442-445.[71] Fages A, Hanghoj K, Khan N, et al. Tracking five millennia of horse management with extensive ancient genome time series [J]. Cell, 2019, 177(6): 1419-1435.[73] Cai D, Zhu S, Gong M, et al. Radiocarbon and genomic evidence for the survival of Equus Sussemionus until the late Holocene [J]. Elife, 2022, 11: e73346.
作者:李畅(吉林大学考古学院);蔡大伟(吉林大学考古学院、吉林大学边疆考古研究中心、吉林大学-西蒙菲莎大学生物考古联合实验室)
原文刊于:《边疆考古研究》第34辑
责编:韩翰