兰州大学最新Nature文章:丹尼索瓦人在青藏高原持续生存到距今约4万年前意味着什么?
发布时间:2024年07月12日
论文摘要:
遗传学和零散的古人类学数据表明,丹尼索瓦人曾经广泛分布在东亚地区。尽管考古证据有限,但这表明丹尼索瓦人能够适应极其多样化的环境。本文我们整合了来自青藏高原白石崖溶洞的中更新世晚期-晚更新世动物群的动物考古学和蛋白质组学分析,在这里发现了丹尼索瓦人的下颌骨和丹尼索瓦人的沉积物线粒体DNA。通过动物考古学分析,我们鉴定了一个新的人类肋骨标本,其年代大约为4.8万至3.2万年前(第3层)。通过高通量蛋白质组学分析,将这个标本分类为丹尼索瓦人血统,将他们在白石崖溶洞的存在扩展到晚更新世。在整个地层序列中,动物群主要由山羊亚科动物组成,还有大型草食动物、食肉动物、小型哺乳动物和鸟类。骨表面上人为改造的高比例表明,丹尼索瓦人是动物群积累的主要因素。对尸体处理的“操作链”表明,动物种类被用于获取它们的肉、骨髓和皮毛,而骨头也被用作制作工具的原材料。我们的结果揭示了丹尼索瓦人的行为以及他们对东亚地区中更新世晚期-晚更新世多样化和不断变化的环境的适应。
原文链接https://www.nature.com/articles/s41586-024-07612-9 科学问题与解决方法 对俄罗斯丹尼索瓦洞穴的几块原始人化石进行的古代DNA分析揭示了欧亚大陆东部尼安德特人的一个姐妹谱系,即所谓的丹尼索瓦人。根据东亚、南亚和东南亚人口中存在的丹尼索瓦人的遗传祖先,可以推断丹尼索瓦人更新世晚期广泛分布在欧亚大陆东部。来自青藏高原东北部甘加盆地白石崖溶洞(以下简称BKC,海拔3280米)的夏河下颌骨(名为夏河1号)和丹尼索瓦沉积线粒体DNA也支持这一主张。它们共同表明,丹尼索瓦人至少从16万年到6万年前左右,甚至可能到4.5 万年前占领了BKC。BKC的考古发掘揭露了一个保存完好的地层,其中包含丰富的石器和动物群,提供了至少约190 ka至约30 ka的原始人类占领的证据。然而,青藏高原上的大多数更新世遗址,如将军府01遗址,只产生了少数零碎的骨骼标本。除了青海湖盆地151个遗址的小型动物群外,青藏高原中更新世和晚更新世没有其他动物考古或古生物学数据。因此,BKC提供了一个独特的机会来研究青藏高原高海拔地区古人类的生存策略及其所处的动物生态学。我们通过液相色谱-串联质谱(LC–MS/MS)分析,建立了青藏高原及其周边地区哺乳动物物种的扩展蛋白质组学参考数据库,以便通过质谱(ZooMS)分析BKC的1857个骨骼和牙齿标本进行高通量动物考古。进一步,我们通过ZooMS将分类学鉴定与来自大量动物遗骸的动物考古数据相结合(n=2567)形成一个重要数据库,这为青藏高原丹尼索瓦人的古生态和生存策略提供了一幅新颖的画面。 白石崖溶洞中动物群落的组成 通过结合形态学和ZooMS鉴定,我们对分析的2567个动物标本中的2005个(78.1%)进行了分类学鉴定。结果显示,羊亚科,主要是岩羊在动物组合中占主导地位。在地层学中,牛科动物的比例很高——例如,羊亚科、野生牦牛和藏羚羊以及马科动物,揭示了在中更新世晚期-晚更新世期间甘加盆地以草本植物为主的景观。森林-灌木物种的存在,如红鹿、麝、槽齿飞鼠和豪猪,反映了小规模的森林-灌木栖息地的存在,类似于现代盆地的山麓和河谷地区。此外,还发现了各种食肉动物(例如,斑鬣狗、狼、藏狐和雪豹以及鸟类(例如,金雕和普通野鸡。 我们在第6层以下鉴定出了灭绝的大型食肉动物和大型食草动物(毛犀牛。虽然较年轻层的样本小于第10层(模拟的最大年龄范围大约是10.9万年到超过22.5万年)和第11层,但目前的数据表明,在形成第6层和第5层(模拟的最大年龄范围大约是6.0-10.4万年)期间BKC周围的动物群落组成发生了显著变化。此外,我们的数据还记录了随着时间的推移羊亚科比例的增加,以及野生牦牛比例的减少。尽管如此,牛科动物和马科动物的持续存在以及在整个地层学中分类多样性的小幅变化表明甘加盆地的环境总体上是稳定的开放环境。 图1:BKC骨表面修饰和骨类型的分布。a、 经分类鉴定的具有食肉动物、啮齿动物或人为修饰的标本的百分比(n=2005)。NISP,已鉴定标本的数量。b、 使用形态学、ZooMS或两者同时使用的不同骨类型中分类鉴定标本的百分比(补充数据5)。骨骼类型基于形态学观察。每个条形图上显示的数字是计算中涉及的相应NISP。c、 每层不同骨骼类型的Caprinae标本的百分比。d、 每层不同骨骼类型上有切割痕迹的Caprinae标本的百分比。e、 具有人为修饰的Caprinae标本的百分比表明每一层的胴体加工活动不同。对于a和c–e,值表示属于相应类别的NISP的百分比。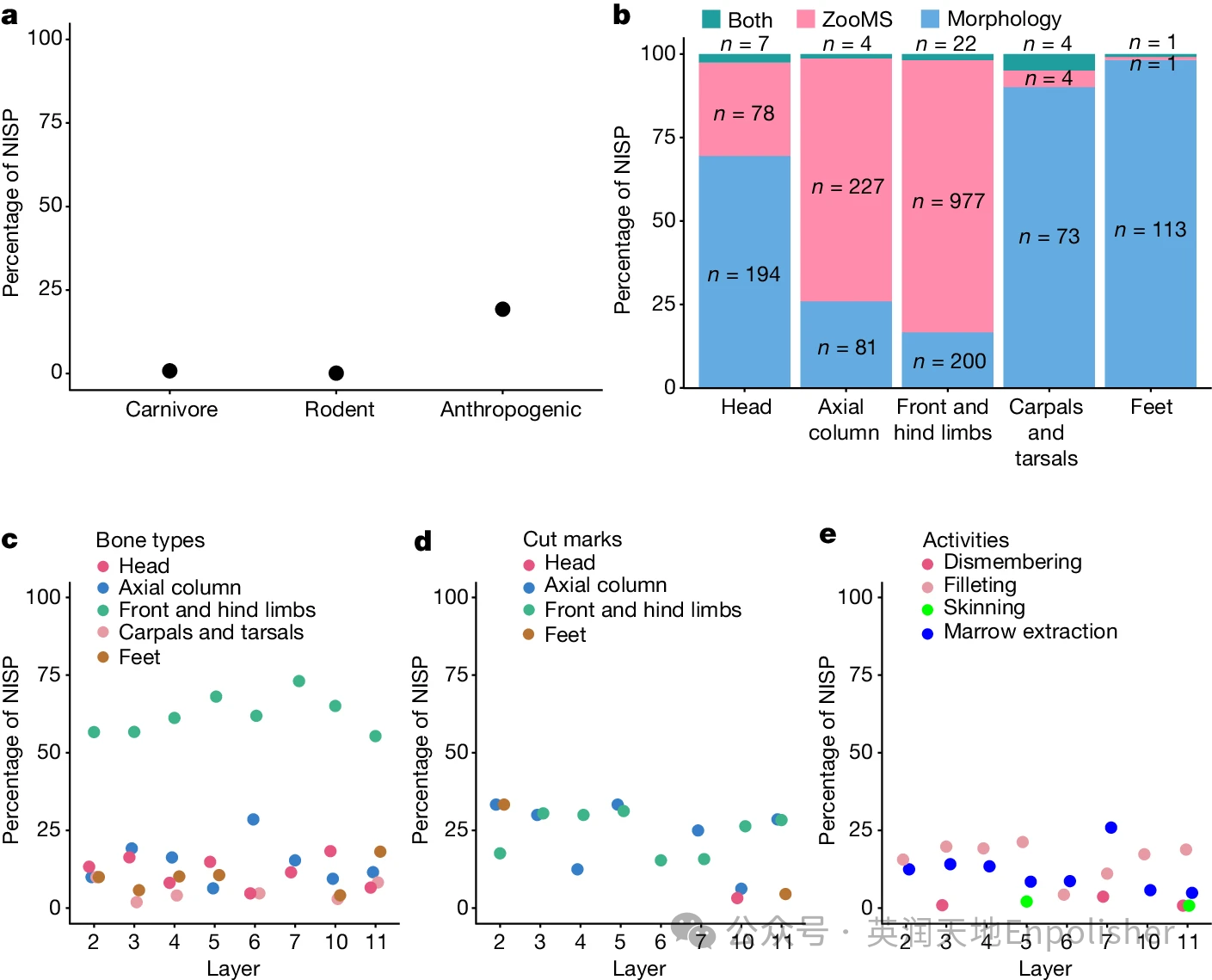
古人类是主要的收集者
BKC的骨骼标本表面保存得非常好,大多数标本(n = 1616,占总数n = 1821的88.7%)处于风化阶段1或2,为近地表风化提供了有限的证据。我们识别出了啮齿类、食肉类和人类活动的痕迹。啮齿类啃咬的痕迹有限(n = 3,占0.1%;图1a),食肉类骨骼被修改的比例略高(n = 16,占0.8%;图1a)。相比之下,更大比例的动物组合显示出人类活动的痕迹(n = 386,占19.3%;图1a)。这些标本大多数是通过ZooMS鉴定的。在草食动物和一些大型肉食动物的骨骼上识别出了切割痕迹和敲击凹槽。人类改造的骨骼比例较高,以及每一层中石器的存在,表明BKC动物组合主要是通过古人类活动积累的。
广泛的人类活动证据
形态学可鉴定的标本主要来源于头部碎片、腕骨、跗骨及相关的足部骨骼(图1b)。ZooMS技术帮助我们能够成功鉴定更多的脊柱和前肢或后肢骨干碎片(图1b)。
在综合数据集中及大多数层中,羊亚科不仅存在于每个层中,而且由所有骨骼部分(颅骨、脊柱、前肢和后肢以及足骨)组成(图1c)。人类活动的程度在时间上与食草动物一样普遍存在。与羊亚科标本相关的切割痕迹是整个地层中最频繁的,除了在第6层和第7层,与骨髓提取相关的敲击凹槽在前肢和后肢骨骼上占主导地位(图1d,e)。除了羊亚科外,其他食草动物的各种骨骼类型上的切割痕迹和敲击凹槽的高频率,表明动物资源的获取并不局限于特定的分类群。
食肉动物、小型哺乳动物和鸟类上也有人类改造的痕迹(图2和扩展数据图5)。在食肉动物标本中多个标本上存在切割痕迹和敲击凹槽(n = 8)(补充表5.2)。总的来说,动物考古学数据表明,BKC的古人类使用了包括大型食草动物在内的广泛物种,以及在较小程度上,食肉动物、小型哺乳动物和鸟类。我们在第11层发现了一种可能的修整器(图2d),并且在第4层、第9层和第10层发现了三件应急骨制工具(图2e)。骨制品似乎来源于在BKC动物组合中占主导地位的物种,而不是故意专注于单一物种。

图2:人类遗传学改良的动物标本和骨骼工具的例子。a,金鹰座右肱骨(第4层),具有浅表和直切标记簇,与羽毛的去除有关。b、 Crocuta Crocuta ultia图谱(第10a层),在断关节过程中产生倾斜切割标记。c、 Marmota sp.(ZooMS分类单元ID)桡骨骨干(第9层),具有由人为破坏产生的负贝壳状髓质片状疤痕(黑色三角形)。d、 一种可能的修饰剂(第11层)。Equus sp.右下P2,颊表面有一组刮痕。e、 方便的骨骼工具(ZooMS分类单元ID:Caprinae;层10b)。该肱骨骨干是通过在其皮质表面上连续直接撞击(由右侧放大图像中的黑色三角形表示)而故意成形的。对于所有面板,放大的图像(a、b、d、e中的右侧和c中的底部)是主图像中用虚线表示的区域的放大率。除非另有说明,分类鉴定均来自形态学分析。比例尺,2厘米(a–d,主图像)、1厘米(e,主图像和1毫米(所有放大图像)。
识别出一个新的丹尼索瓦个体
在对无法识别的碎片进行ZooMS筛查过程中,一个肋骨标本被鉴定为人亚科(图3a和扩展数据图6a)。该标本含有14个与人亚科匹配的I型胶原蛋白(COL1)肽标记,以及一个仅在Hominoidae中发现的肽标记(补充数据4和扩展数据图6a)。考虑到其他大型猿类当前和过去的地理分布,特别是Pan属,这个标本可以被确认为现代人。因此,我们将这个古人类标本命名为夏河2。夏河2标本来自T3的第3层,其年代已被定为48-32ka。夏河2的谷氨酰胺脱酰胺值(酸解COL1α1 508-519 = 0.52和酸解COL1α1 435-453 = 0.46)与第3层的其他标本以及直接放射性碳定年到大约50-30千年前的标本相似(扩展数据图6b)。进一步的鸟枪法蛋白质组学分析为夏河2的分类归属提供了更具体的信息。夏河2始终与已发表的高覆盖度丹尼索瓦人基因组聚在一起(图3b)。因此,可以确定,在现有的参考个体中,夏河2个体与D3丹尼索瓦人高覆盖度个体关系最为密切。夏河2标本的拓扑结构和位置与之前对夏河1下颌骨分析得到的结果相似。夏河2丹尼索瓦人的发现将在白石崖洞中丹尼索瓦人存在的化石证据从中更新世晚期一直延伸到晚更新世,与从BKC第3层回收的丹尼索瓦沉积物线粒体DNA一致。
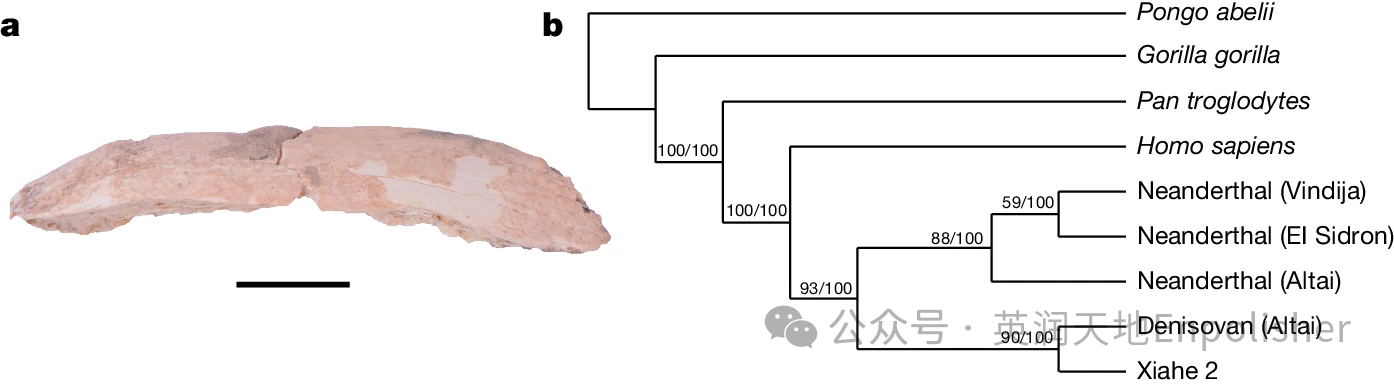
图3:夏河2号标本,一个通过动物质谱筛查发现的人属肋骨标本。比例尺,1cm.b,夏河2号标本和参考蛋白质组的系统发育树。分别针对最大似然和贝叶斯分析显示了节点处的支持值。
讨论与结论:
先前的研究表明,BKC(白石崖溶洞)是目前已知的青藏高原上唯一保存完好的洞穴遗址,跨越了中更新世晚期至晚更新世。夏河1下颌骨和沉积物线粒体DNA(mtDNA)分析显示,丹尼索瓦人至少在约16万年、10万年(第7层)和6万年(第4层)占领了这个洞穴,并且可能一直持续到大约4.5万年(第4层末端)。在这里确认的夏河2肋骨,以及从第3层发现的丹尼索瓦沉积物mtDNA,表明丹尼索瓦人至少在4.8万至3.2万年前占领了这个地点。此外,夏河1的最小铀系年代约为16万年,这也与第10层或以下的时间年代相符(图4g)。尽管这并不能证明夏河1肯定来自BKC的第10层、第11层或更老的层,但它确实表明,在这些层形成期间,夏河丹尼索瓦人是最有可能的居住者。
到目前为止,BKC的第3至11层还没有发现其他古人类的证据,同样,在同期青藏高原东北部也没有发现其他古人类居住的证据。因此,合理推测丹尼索瓦人至少从大约16.7万年前占领了BKC,直到大约4万年前,可能从超过22.4万年前(第10层的模拟最大年龄)到3.2万年前。因此,BKC的动物群组合记录了丹尼索瓦人在甘加盆地最后一次冰期-间冰期-冰期周期中的行为和生存方式(图4)。

图4:BKC的区域和北半球气候历史、动物群生态和丹尼索瓦人的占领。a、 LR04海底堆栈δ18O记录。b、 10中国北部宝鸡黄土样品的Be基降雨。c、 中国黄土高原的磁化率(归一化)记录。d、 西藏高原东缘若尔盖盆地的乔木花粉(AP)百分比(约600年分辨率)。e、 BKC地层单元的Shannon指数。f、 来自骨骼标本的肽COL1α1 508–519的脱酰胺值。紫色点和误差条表示每层骨骼的平均去酰胺值,概率范围为68.2%。蓝色三角形表示具有放射性碳年代的2-3层单个骨骼样本的脱酰胺值(n=9,补充数据2)。绿色的心表示两个丹尼索瓦人标本(夏河1号和2号)的脱酰胺值。g、 地层和丹尼索瓦沉积mtDNA。从2、3、4和7层沉积物中提取的丹尼索瓦线粒体DNA由绿色恒星表示。h、 各层的建模年龄范围。黑点和条形范围表示各层的建模平均年龄和年龄范围(补充表2.2)。第2层的年龄范围仍在评估中,目前还没有第5层和第11层的日期。层5的指示年龄范围估计基于层4和层6之间的年龄间隔。
对BKC动物群组合的全面分析显示,丹尼索瓦人利用了甘加盆地周围以草本植物为主的景观中存在的广泛动物类群。Caprinae(羊亚科)在古人类居住期间,尤其是在MIS 5之后的第5至第2层,变得越来越突出,其中Caprinae标本占动物群组合的一半以上。对Caprinae标本的人为改造分析表明,包括系统屠宰和使用骨骼材料制作工具在内的完整加工流程在BKC是存在的。除了Caprinae,大型草食动物、食肉动物、小型哺乳动物和鸟类的遗骸也以多种方式被使用。这表明丹尼索瓦人充分利用了他们可用的动物资源,以在最后一次冰期-间冰期-冰期周期中的高海拔青藏高原上生存。在冰期和间冰期,甘加盆地可能提供了一个适宜的避难所,尽管海拔较高,但资源相对稳定,特别是与青藏高原的更高海拔地区或邻近的中国黄土高原上波动的环境条件相比。
通过将BKC与其他丹尼索瓦人或可能的丹尼索瓦人遗址进行比较,即俄罗斯的丹尼索瓦洞穴和老挝的眼镜蛇洞洞穴,我们发现它们的动物群组合与它们各自的地理环境相匹配,对应于高海拔、高纬度和热带(或亚热带)环境。因此,我们的结果为丹尼索瓦人的古生态和行为可塑性提供了证据。此外,这些见解引发了关于丹尼索瓦人在青藏高原灭绝的原因和时间、以及丹尼索瓦遗传特征在现代人类中的起源的问题。
文章来源:“英润天地Enpolisher”公众号