浙江大学方盛国教授牵头开展大规模基因组研究改善了对大熊猫种群结构的理解并提出未来的保护措施
发布时间:2024年09月13日

建立保护区是保护小型种群,尤其是濒危物种的重要策略之一。划定物种的精细遗传结构有助于合理规划保护区并保持遗传多样性。如果遗传结构反映了祖先遗留的变异,在进行物种从一个区域到另一个区域的迁移时,考虑异种交配风险非常重要,因为在长时间进化历史中形成的地方适应性在不同种群间可能有所不同。相反,如果某物种的遗传结构是由于栖息地破碎导致的隔离所致,种群间的基因流动可能更好地减少遗传多样性的丧失。
遗传拯救被认为是一种通过防止近亲繁殖和促进基因流动来增加遗传多样性、提高适应性的重要手段。因此,了解特定物种的每个种群的基本遗传背景和灭绝风险是制定基于证据的遗传拯救计划的关键,这包括整个基因组的遗传多样性、遗传分化、历史和当前的基因流动、近亲繁殖以及潜在的有害突变。
大熊猫(Ailuropoda melanoleuca)是全球生物多样性保护的旗舰物种,被划分为两个亚种:仅分布在秦岭的秦岭大熊猫(于1960年发现,1964年北京师范大学郑光美首次提出,浙江大学生命科学学院方盛国的研究小组于2005年通过形态学以及分子生物学方法证实)和生活在岷山、邛崃山、大相岭、小相岭和凉山的四川大熊猫。生态和生物因素的挑战将大熊猫推向了濒危状态。中国政府在过去几十年中通过了一系列法律以加强对野生大熊猫种群的保护。根据2014年发布的第四次全国大熊猫分析报告,野生大熊猫数量从1999-2003年的1596只增加到2011-2014年的1864只,因此大熊猫在2016年被国际自然保护联盟(IUCN)从“濒危”降级为“易危”。
然而,由于基础设施、农业和旅游业的发展,当前人类活动带来的威胁比几十年前更加严重,这些发展大多发生在大熊猫的原始栖息地。正如第四次全国大熊猫普查的那样,野生大熊猫种群被分成33个孤立的亚种群,其中18个亚种群的个体数量少于10只。显然,尽管大熊猫在近期内不太可能灭绝,但一些野生种群仍面临局部灭绝的风险。在过去的二十年中,通过在孤立种群之间建立生态走廊和将代表野生谱系的圈养大熊猫重新引入野外,这些保护策略已经得到实施并取得初步成功。2021年,中国政府宣布开设跨越四川、陕西和甘肃省的国家大熊猫公园,进一步扩大了对大熊猫的保护。此外,分子证据和基因组分析揭示了秦岭亚种以及四川大熊猫的初步种群结构。然而,关于野生和圈养大熊猫种群的基因组背景仍大多未知,这些信息对于深入了解它们的遗传状态、帮助未来的保护工作至关重要。
最近,关于濒危物种的全面基因组背景研究已被报道用于协助保护工作。同样,对于大熊猫来说,系统的种群基因组分析对于确定详细的种群结构、进化历史和灭绝风险非常重要,这些都是制定短期和长期保护计划的关键。在本研究中,作者通过开展大规模的种群基因组学研究,涉及649只大熊猫,覆盖六个山地种群和圈养个体,全面分析和比较了野生和圈养大熊猫的遗传背景,作者认为这些研究将有助于未来基于证据的大熊猫保护工作。
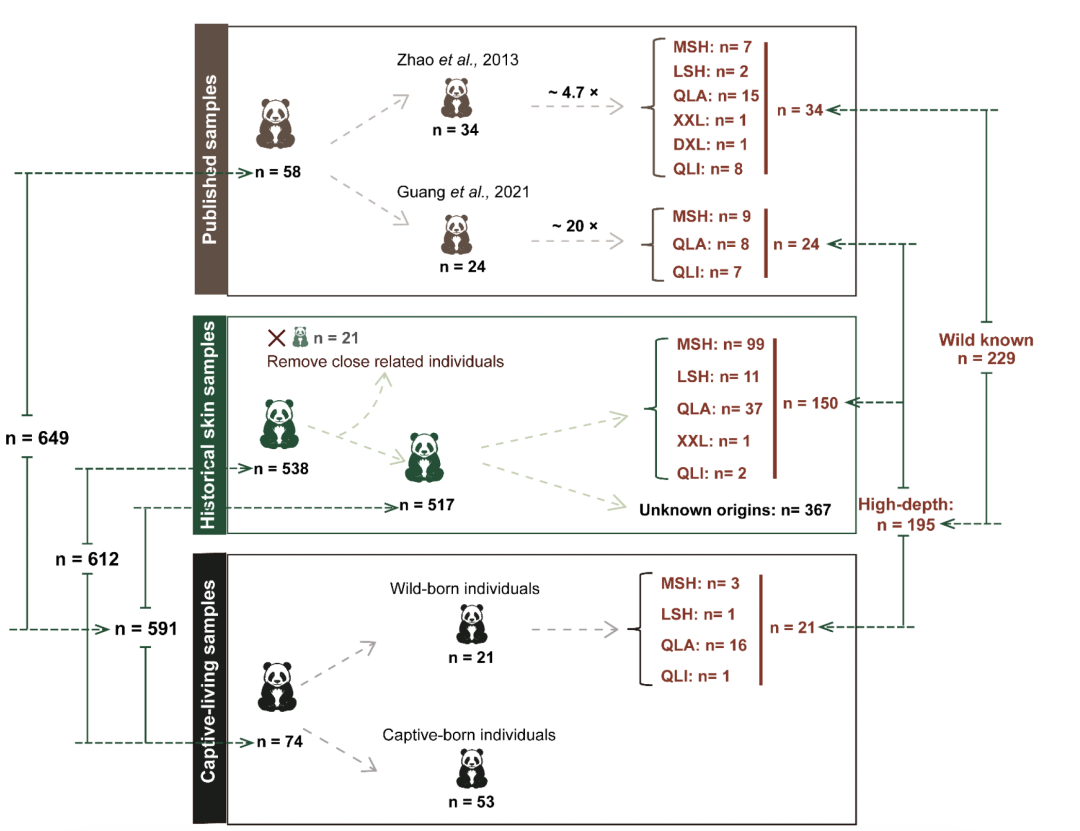
样本和基因组测序
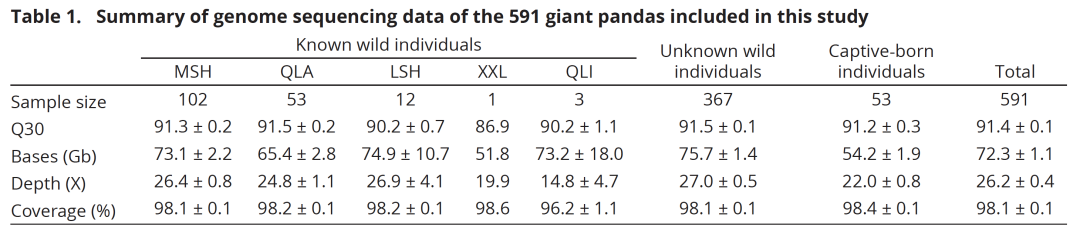
对612只大熊猫进行了全基因组重测序,包括538只野生个体和74只圈养个体,平均测序深度为26.08 ± 0.34,覆盖率为98.10 ± 0.03%。考虑到一些来自野生大熊猫的旧皮肤样本缺乏详细信息,首先进行了物种鉴定和个体化分析,以便于后续的基因组分析。发现所有样本均来自大熊猫,但其中39个样本之间存在密切亲缘关系,因此随机从每对样本中选择一个,最终保留了517只无亲缘关系的野生个体,其中150个样本具有已知的采样地点。
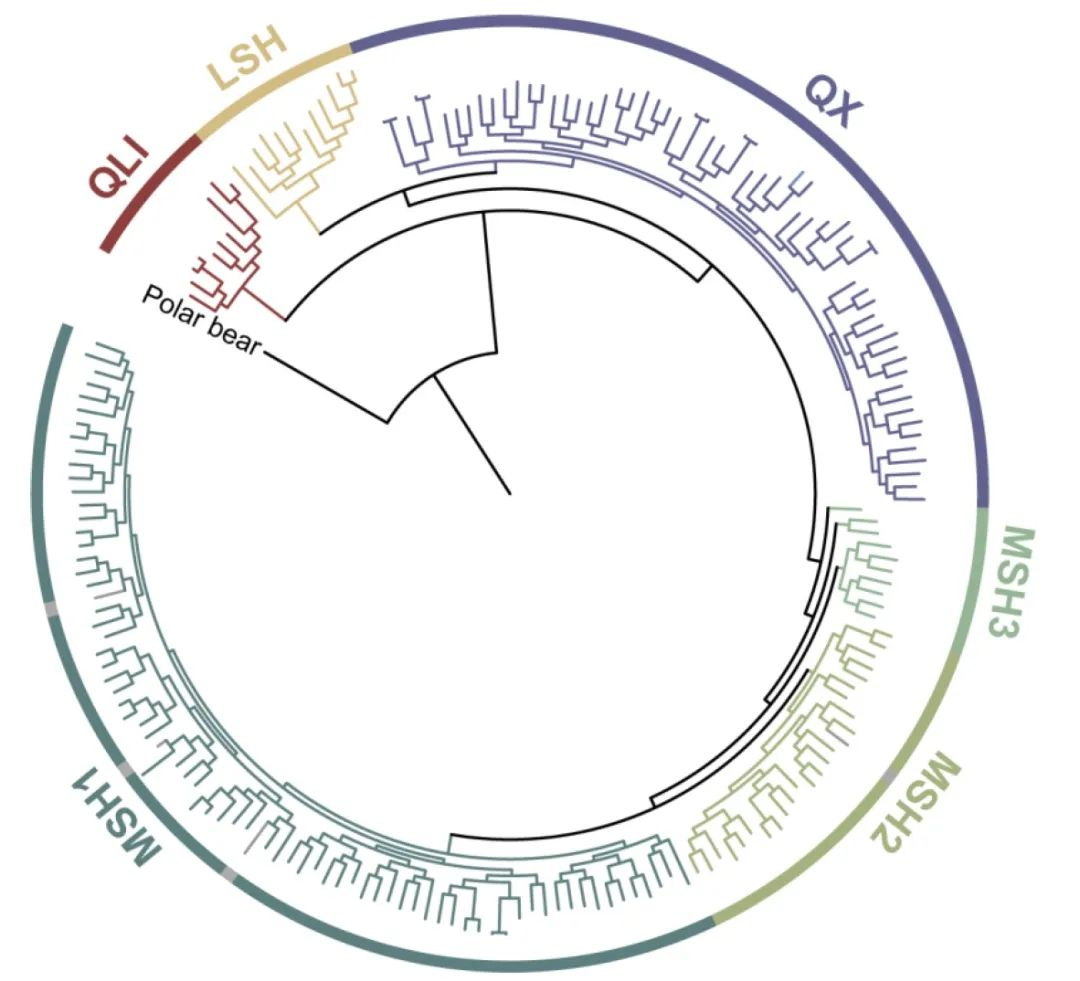
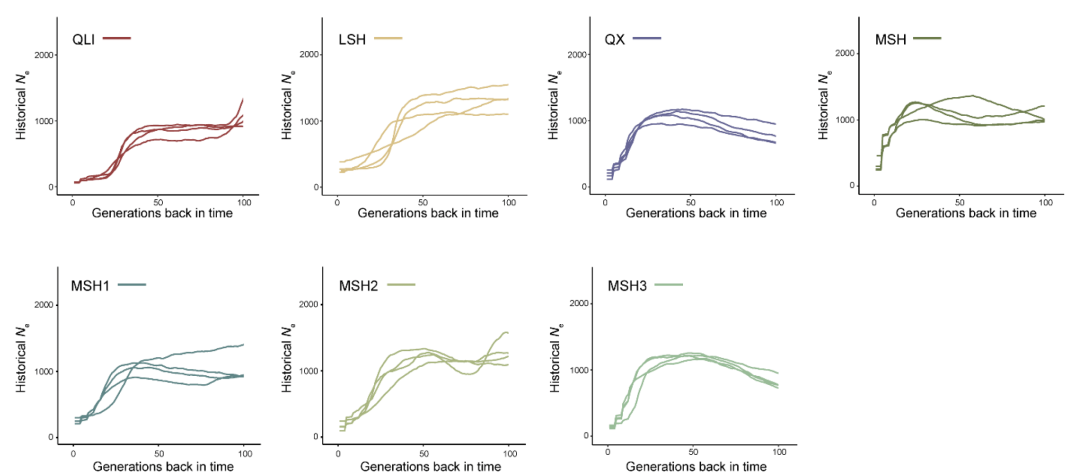
仔细检查PCA聚类时,注意到岷山种群由三个亚群组成(MSH1,MSH2,MSH3)(图2A)。混合分析也支持在K=6时岷山种群分为三个聚类(图2C),最大似然树显示了相应的三个分支,但还能鉴定出更多的亚分支(图2B),揭示了岷山种群的复杂结构。进一步分析了仅来自岷山的样本,结果与鉴定出三个独立亚群的结果几乎相同(图2D和E)。有趣的是,除了四个样本外,这三个亚群与它们的地理分布相吻合,MSH1、MSH2和MSH3分别对应岷山的G、J和K区(图1和图2D)。
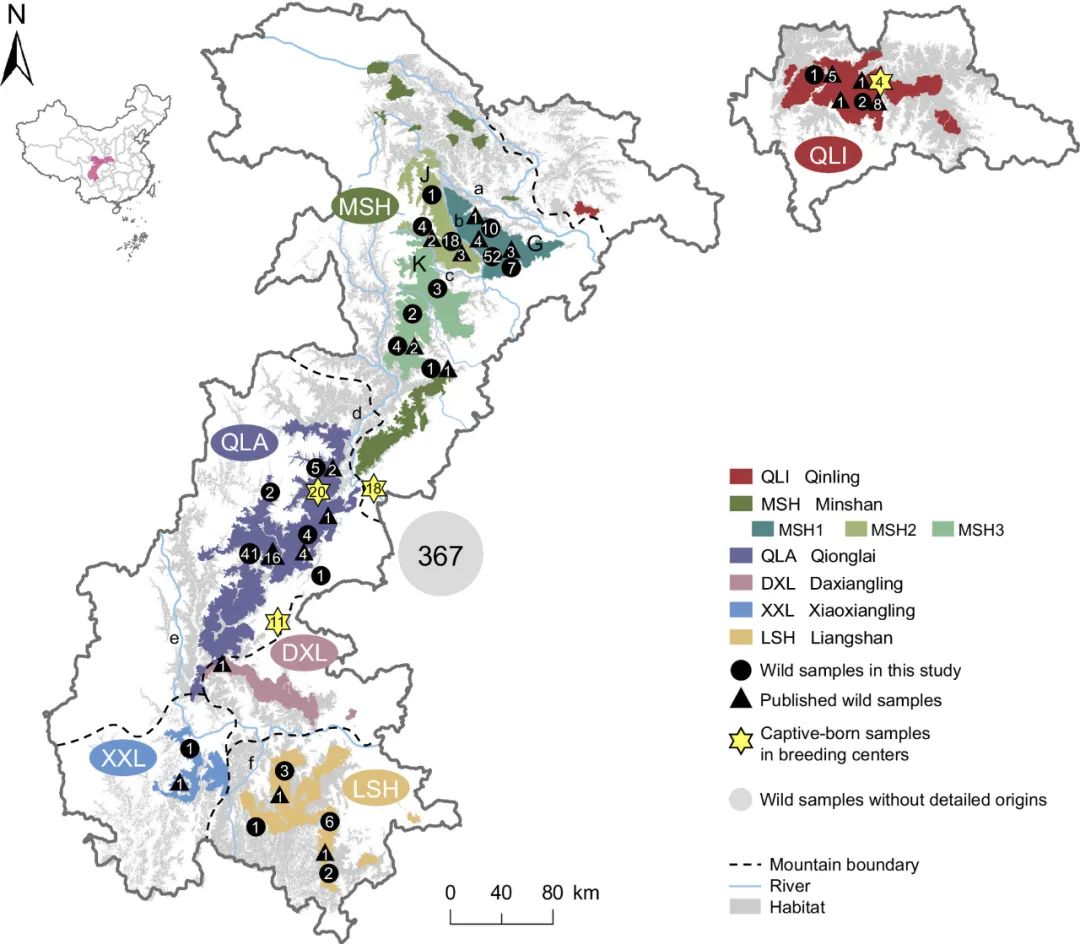
图1 | 本研究用到的大熊猫采样信息
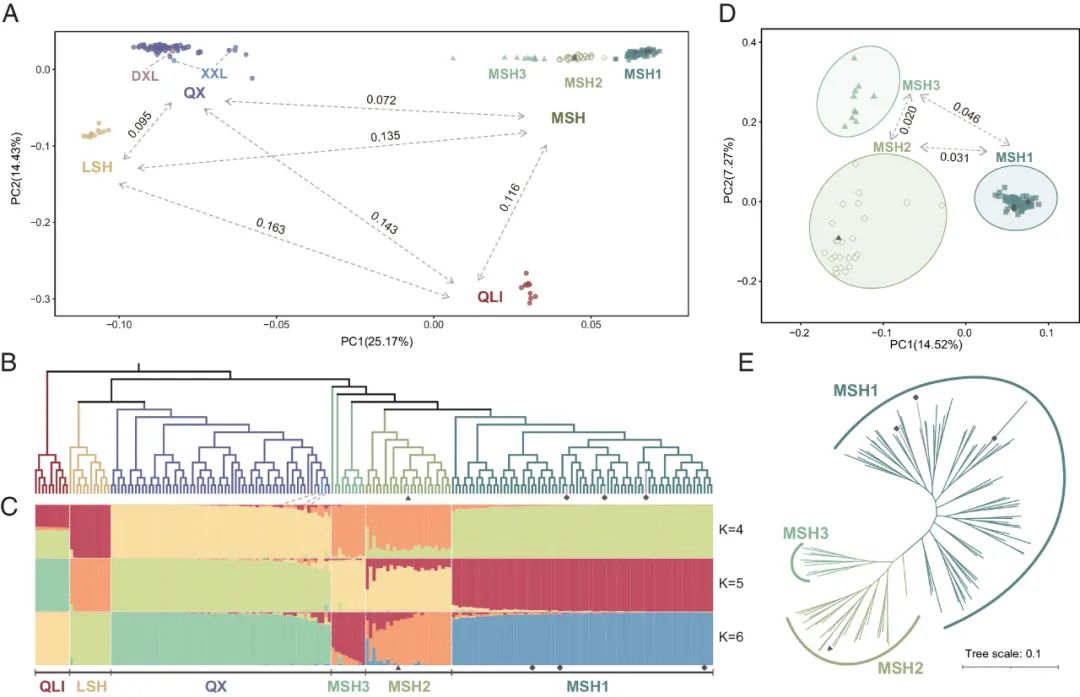
图2 | 野生大熊猫种群遗传结构
基于这一遗传结构,尝试使用监督和无监督聚类算法将其余367个采样信息未知的个体分配到当前的大熊猫种群中。最终,鉴定出256个个体最有可能来源于岷山种群,16个个体最有可能来源于凉山种群。其余95个个体无法明确分配到某一独立种群,因为这些个体与邛崃山、大相岭和小相岭种群相关,支持上述提到的QX种群。通过将这367个样本添加到分析中,再次证实了岷山种群中存在三个亚群。这些关于种群结构的信息对于未来野生种群的管理具有重要价值,但应通过更多基因组数据,特别是来自小相岭和大相岭的数据,进一步证实。
野生种群的历史基因流和混合
系统发育分析支持秦岭种群与其他四川种群最早的分化,但这些种群的历史基因流和混合事件需要进一步阐明。我们首先通过结合系统发育树、同源片段(IBD)和TreeMix分析中的证据,检查了进化拓扑结构((((MSH, QX), LSH), QLI))。基于这一拓扑结构,我们利用北极熊作为外群,理清了种群之间的基因流。
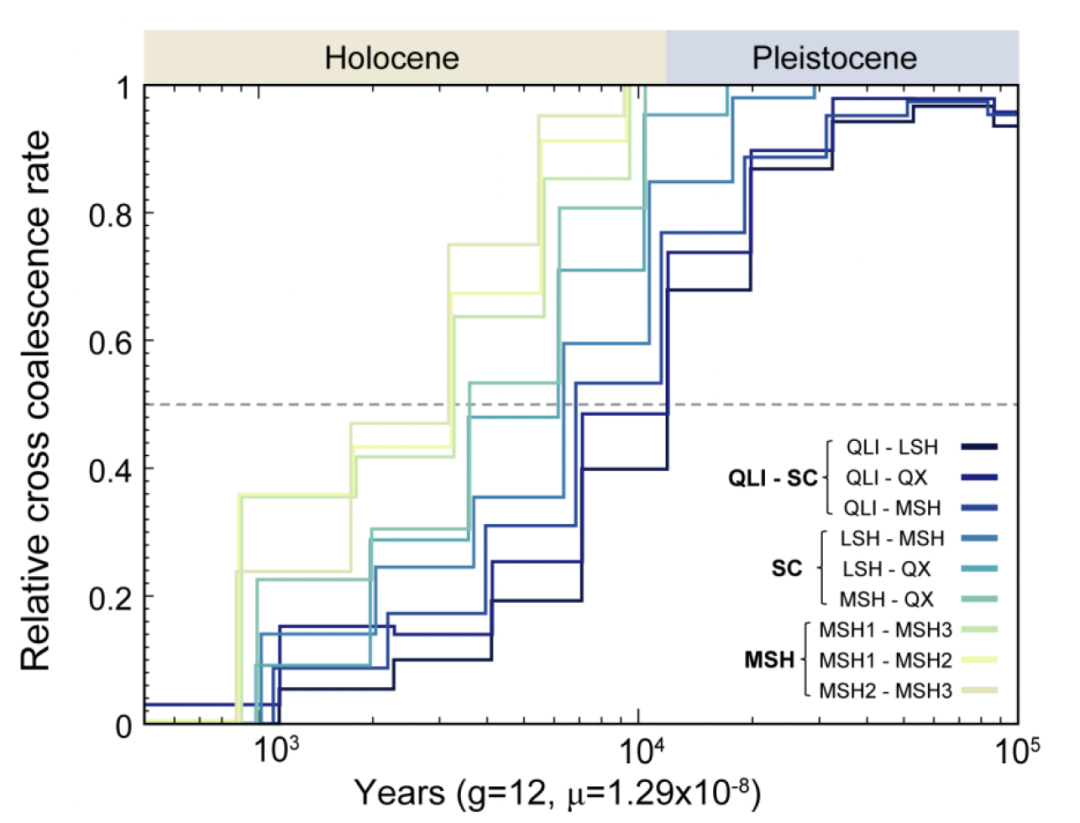
发现最强的基因流发生在秦岭和岷山种群之间,这一发现得到了D统计量、TreeMix分析以及秦岭和岷山种群之间线粒体单倍型共享的支持。特别是,秦岭和MSH1亚群之间的基因流动比秦岭和MSH2或MSH3亚群之间更为频繁。在四川种群中,凉山和QX种群之间的基因流动比凉山和岷山种群之间更为频繁。有趣的是,QX与MSH3亚群之间的基因流动比与MSH1和MSH2亚群之间的更为频繁,这与它们的地理分布一致(图1)。尽管IBD分析显示出自2到3万年前以来大熊猫种群之间频繁的基因流动,但在最近的1000年内,秦岭与四川种群之间以及凉山与岷山种群之间的基因流动已经减少到几乎不存在的程度。
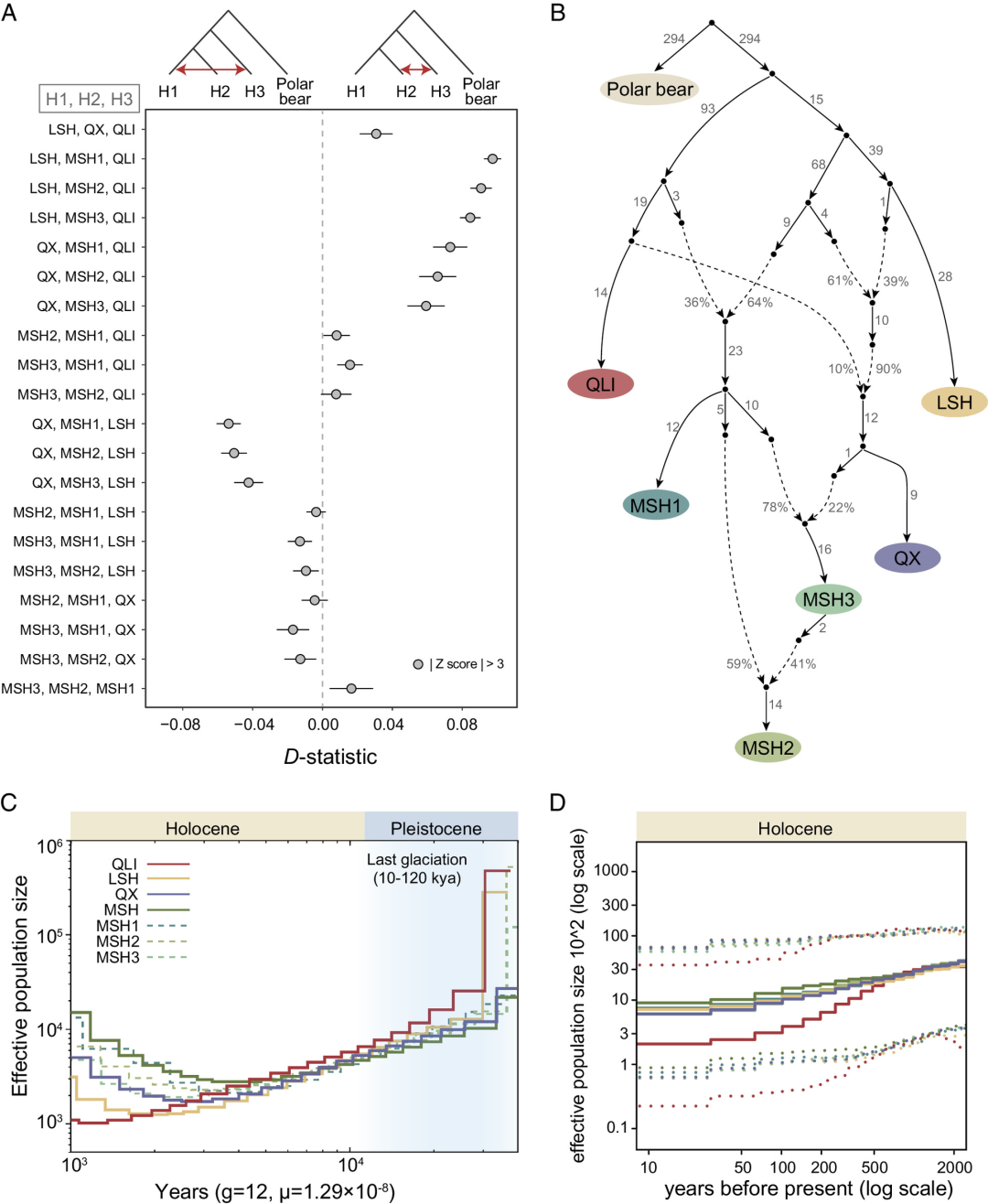
图3 | 现存野生大熊猫的近期种群历史
通过qpGraph建模推断出的混合场景全景非常复杂(图3B)。秦岭种群的一个古老谱系迁移到了岷山种群的祖先。在岷山种群几乎同时分化为三个分支后,QX种群的遗传成分进入了MSH3亚群,然后又贡献给了MSH2亚群。因此,岷山和QX种群的形成涉及与秦岭和凉山种群的古代杂交事件,为恢复过去的基因流提供了进化基础。
大熊猫的种群历史
推测大约在12千年前(kya)至7千年前,秦岭大熊猫种群(QLI)从四川大熊猫种群中分离出来,随后在四川大熊猫种群中最早发生分化的是LSH。MSH种群和QX种群之间的分化发生在6千年前至3.5千年前,这与之前的估算一致。
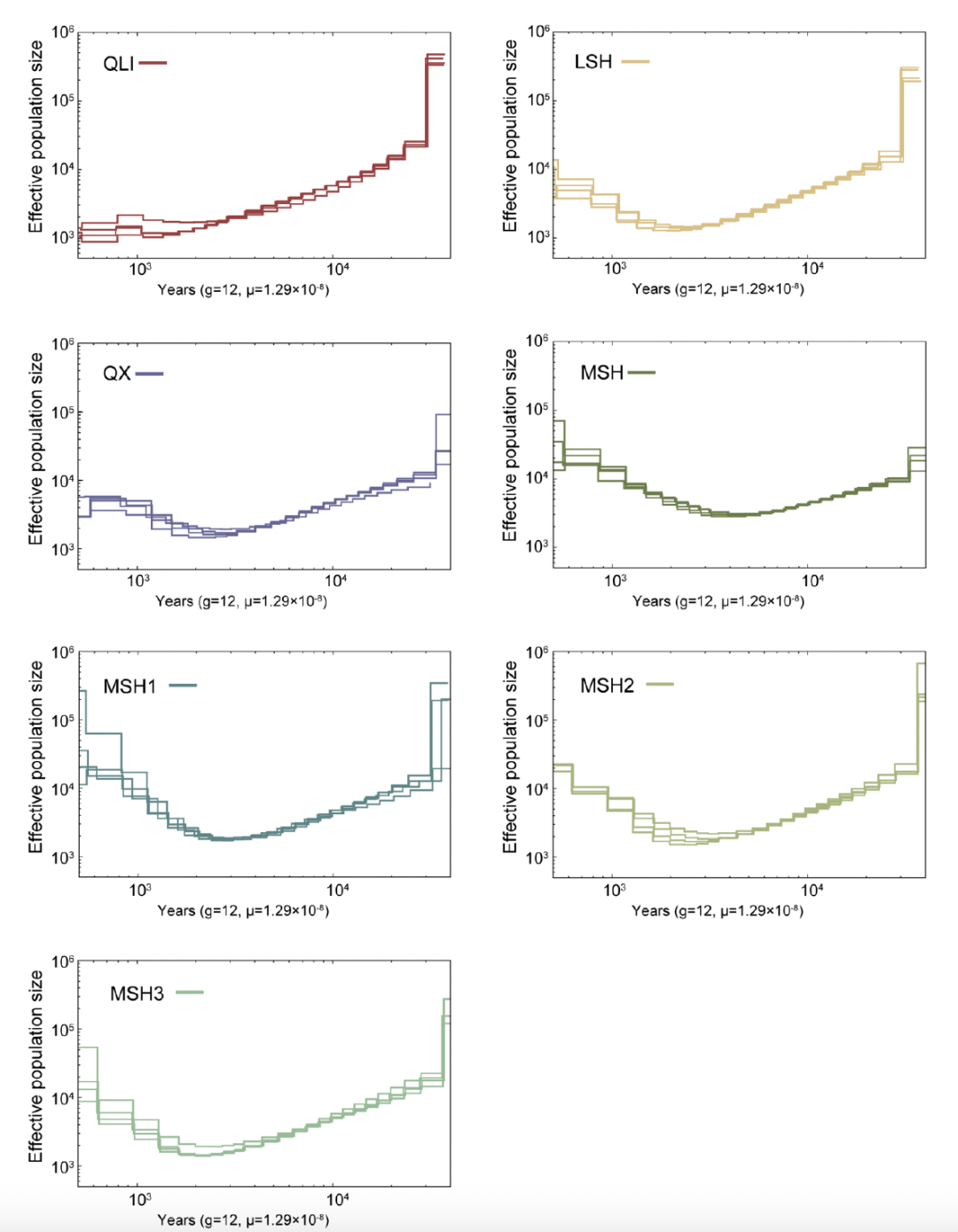
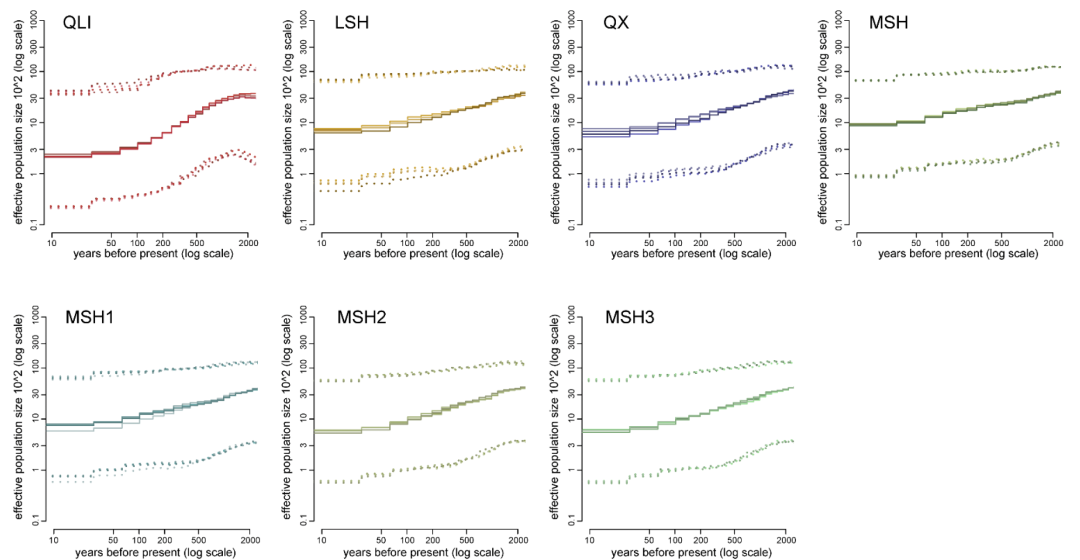
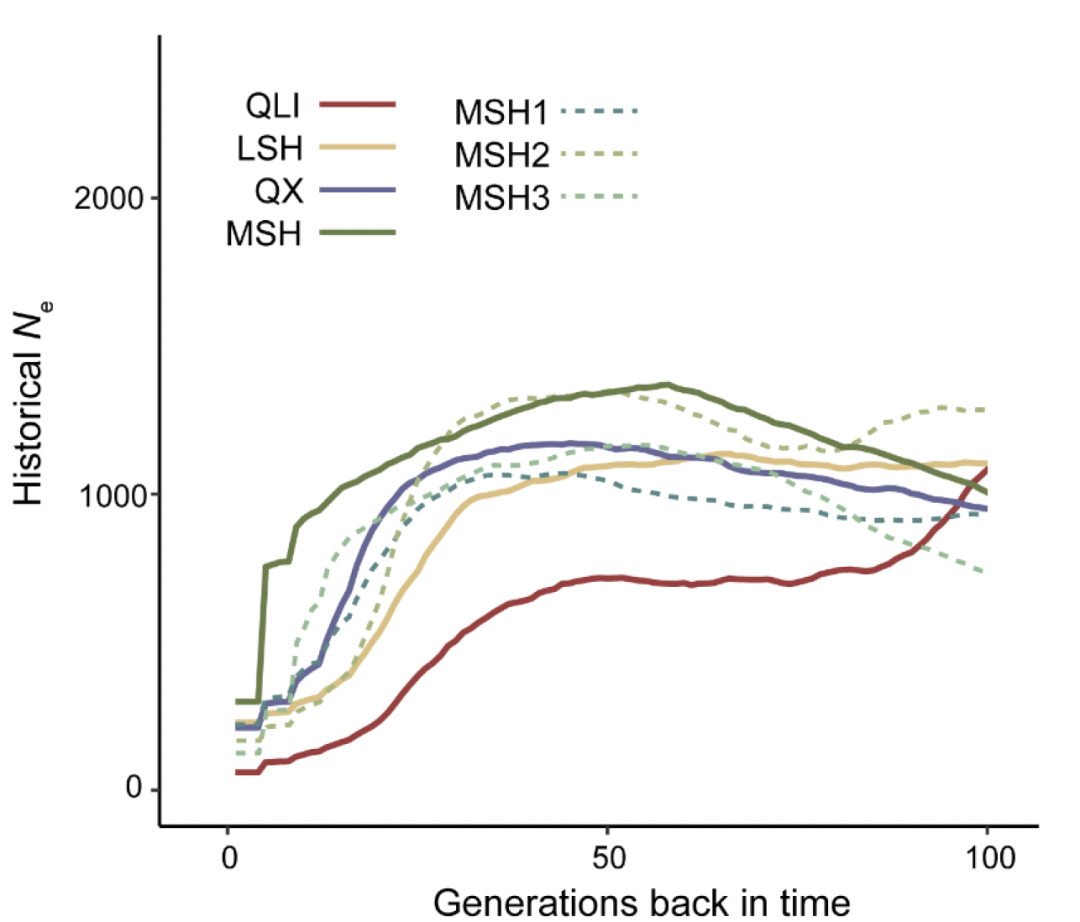
接着,作者揭示了秦岭和四川亚种分化后的种群轨迹。三个四川种群(MSH、LSH和QX)经历了相似的种群历史,从3万年前到1万年前经历了持续的下降,随后在过去1万年中经历了不同的有效种群大小( Ne)变化(图3C)。更严重的是,与四川种群相比,秦岭种群从3万年前到现在快速下降,没有出现种群反弹(图3C和3D)。然而,秦岭种群的Ne 在7千年前之前最大,然后与四川种群的Ne 趋同,并一直保持在最低水平至今。
大熊猫种群的近亲繁殖情况
通过全基因组鉴定纯合片段(ROH),评估了大熊猫种群的近亲繁殖情况。在野生大熊猫中,所有ROH片段的平均长度和最长长度分别为316.23 Kb和8.79 Mb;而在圈养大熊猫(CBC)个体中,分别为332.52 Kb和10.89 Mb。在野生和圈养大熊猫中,长度小于1 Mb的ROH占主导地位(野生:95.98%;圈养:94.85%),表明近期近亲繁殖不严重。检测到野生和圈养大熊猫的ROH比例( )分别为23.52 ± 3.79%和18.30 ± 6.56%(图4A),表明圈养大熊猫面临的近亲繁殖风险显著低于野生大熊猫。作者还发现了不同染色体上ROH长度和数量的分布模式。
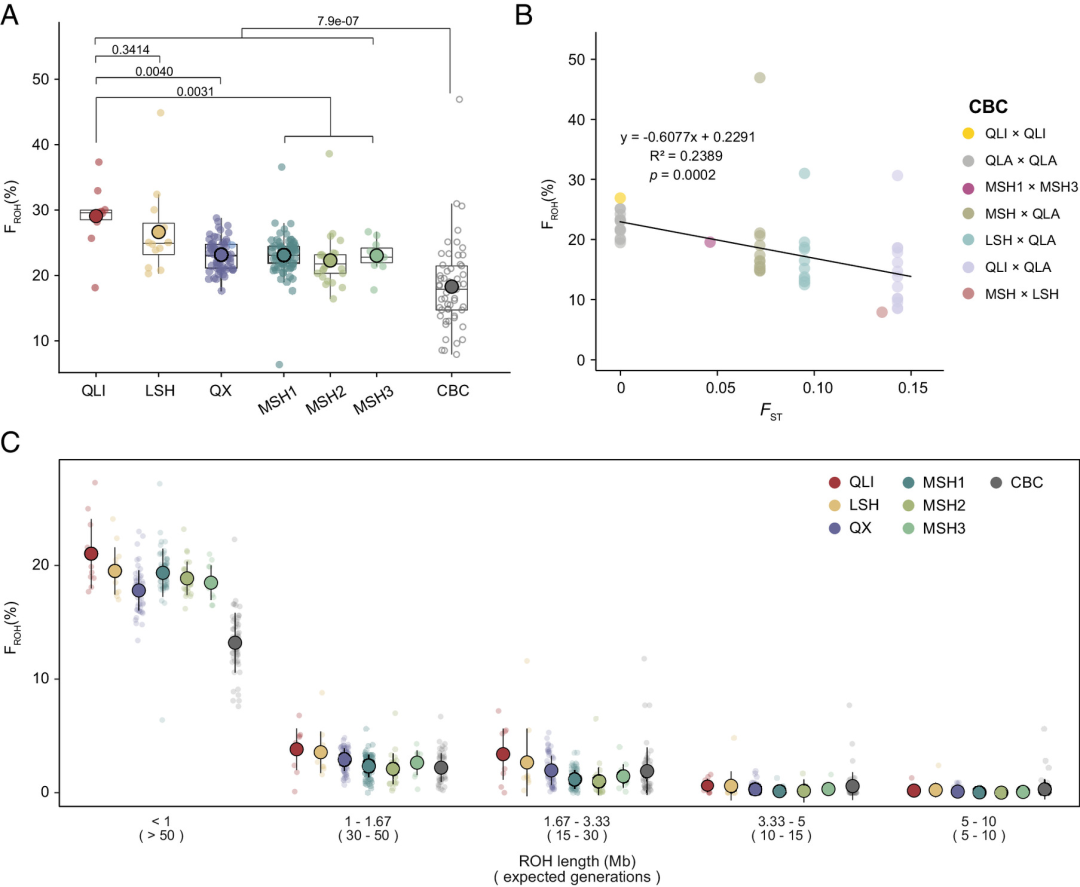
图4 | 野生和圈养大熊猫种群近交水平

进一步分析了各个野生和圈养种群中的近亲繁殖水平。我们发现秦岭种群拥有最多的ROH,长于其他种群。如预期,秦岭种群的 高于四川种群。同样,秦岭种群的遗传多样性( )也是所有野生大熊猫种群中最低的。对于圈养个体,值得注意的是,杂交圈养大熊猫(hCBC,父母来自不同野生种群的祖先)的近亲繁殖水平远低于近亲繁殖圈养大熊猫(iCBC,父母来自同一野生种群的祖先),显示了通过人工交配管理减少近亲繁殖的价值。此外,圈养大熊猫的 与父系和母系血统之间的遗传分化呈负相关(图4B)。
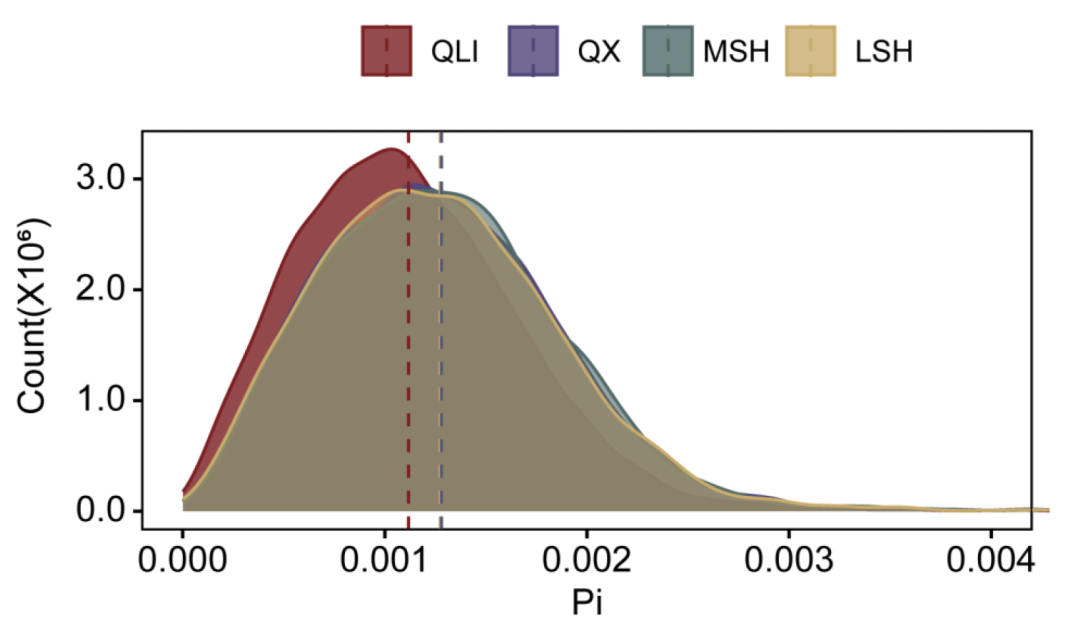
大熊猫的遗传负荷
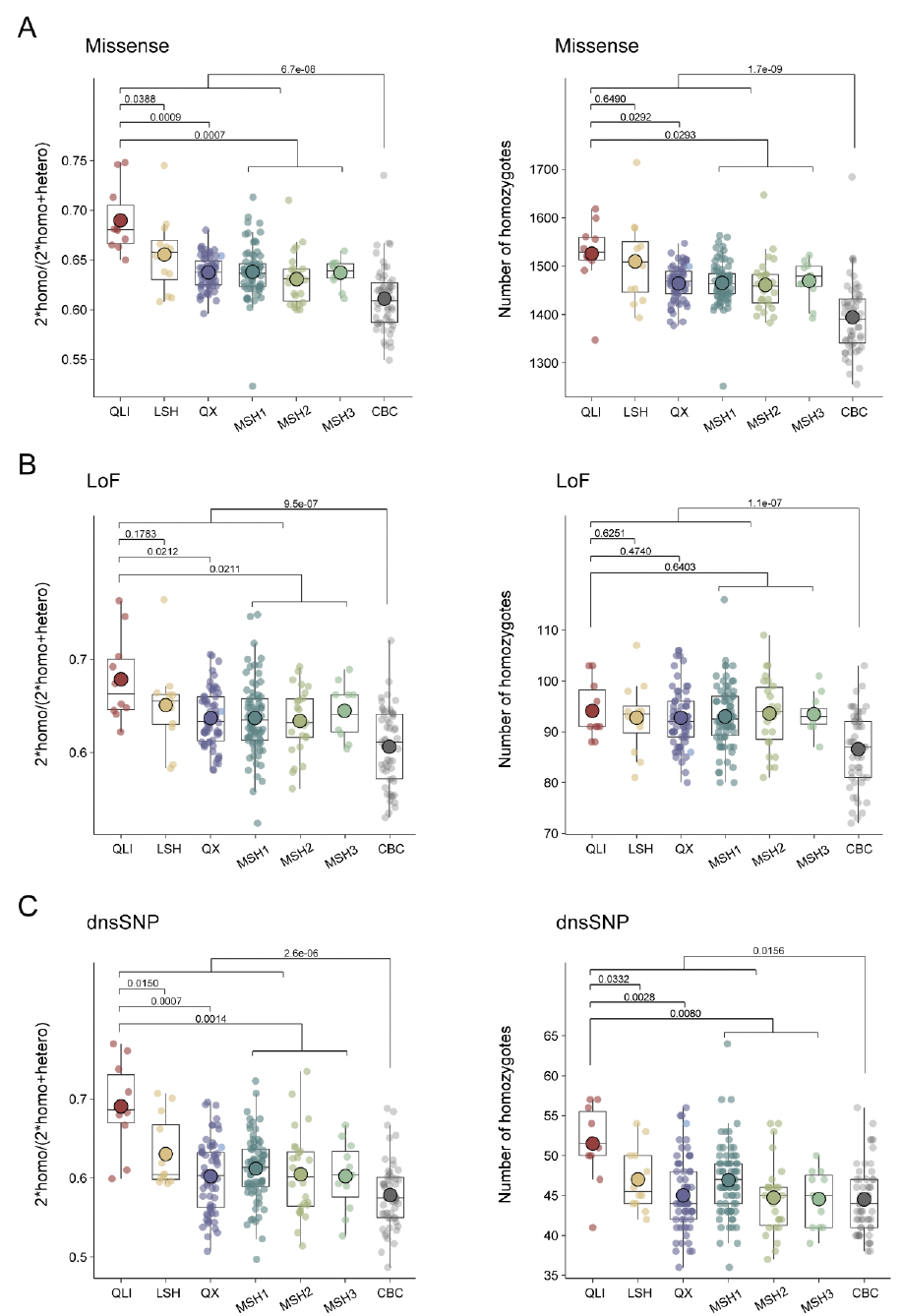
通过全基因组筛查错义突变、功能丧失(LoF)突变和有害的非同义SNP(dnsSNP)以鉴定有害突变。不出所料,秦岭种群的有害纯合突变最多,其比例显著高于三个四川种群(图5A)。进一步发现,所有野生种群中,纯合LoF与 呈正相关(图5B)。发现许多高频LoF分布在与免疫系统和生殖相关的基因中,其中秦岭种群在与生殖相关的基因中携带最多的LoF,而凉山种群在与免疫相关的基因中携带最多的LoF。
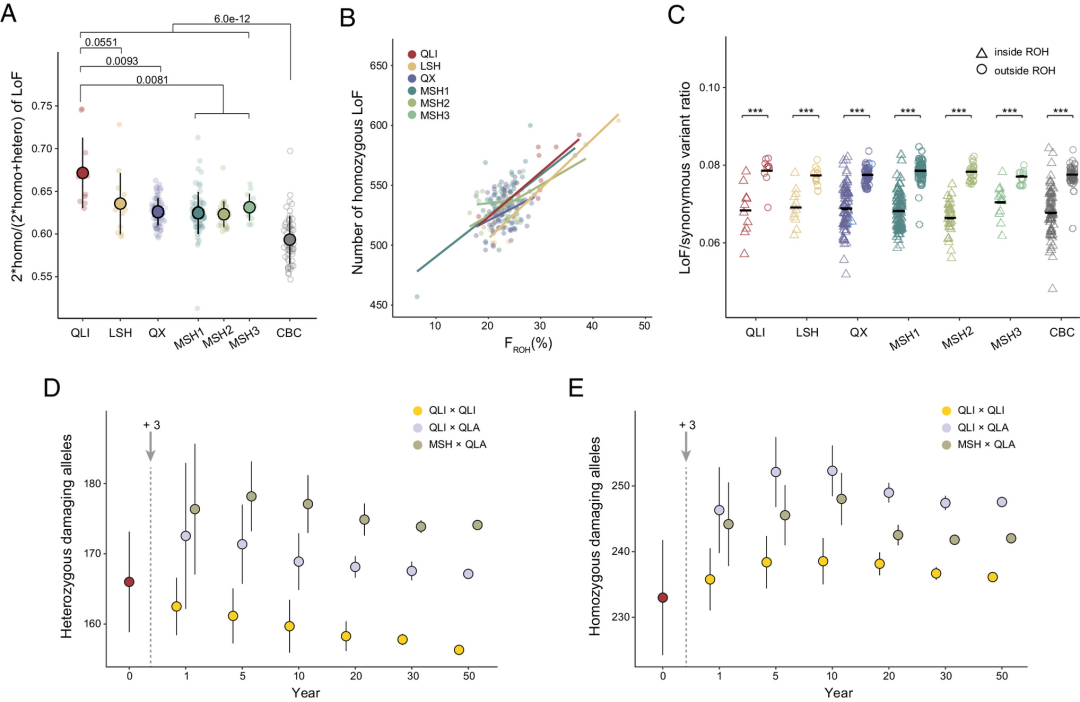
图5 | 大熊猫种群的遗传负荷与野化放归模拟
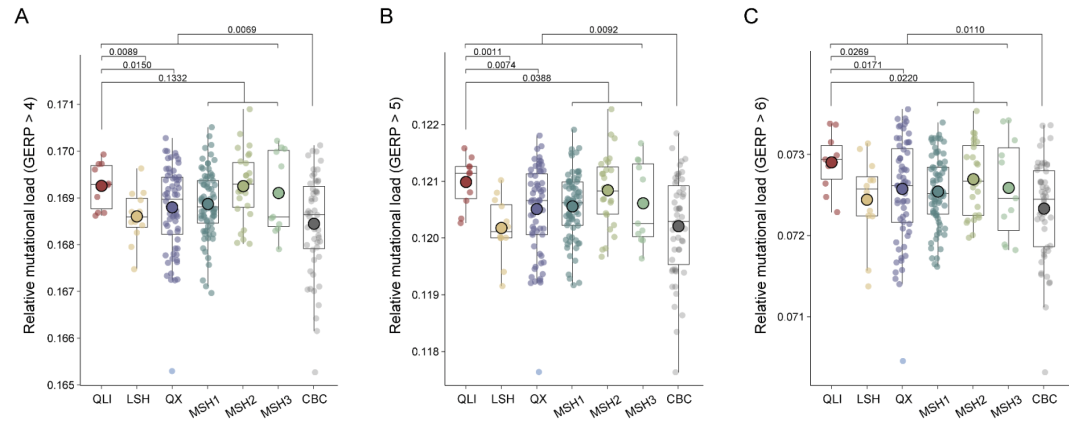
圈养大熊猫中的有害纯合突变少于野生大熊猫。还发现圈养个体中的LoF和 之间存在显著的正相关。有趣的是,这种相关性在近亲繁殖圈养大熊猫个体(iCBC)中比在杂交圈养大熊猫个体(hCBC)中更强,这表明通过交配管理,未来几代可以进一步缓解遗传负荷。为了证实这些发现,进行了GERP分析,以鉴定全基因组中最保守的位点,提供了潜在最有害突变的信息。分析结果与SnpEff预测的结果一致。
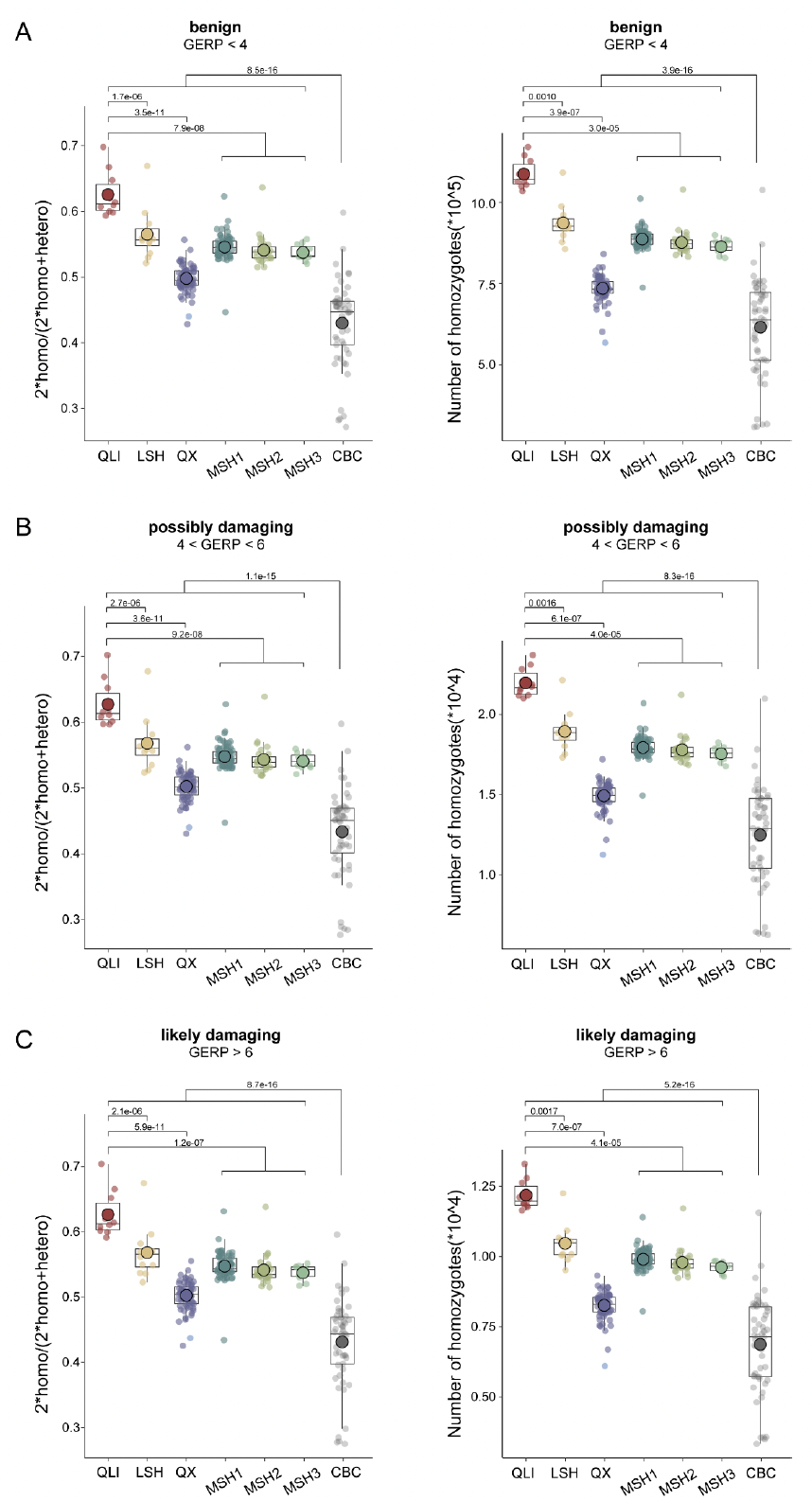
此外,将携带有害等位基因的基因与人类基因突变数据库(HGMD)进行了比对,以缩小可能与病理性疾病相关的突变范围。HGMD筛查显示,秦岭种群的有害遗传负荷最高,其次是凉山种群。再次,圈养大熊猫的总体有害遗传负荷低于野生大熊猫。
遗传清除
大效应的有害等位基因通常是隐性的,并在大种群中被掩盖,但在小种群中暴露时可能导致近亲繁殖衰退。遗传清除经常发生,但在小种群中更为常见,通过近亲繁殖减少影响适应性和存活能力的纯合形式的大效应有害突变,这种清除过程在纯化选择下更加有效。
由于本研究中采样的所有大熊猫个体均为成熟的幼年或成年个体,因此严重影响生存的有害突变在ROH区域中以纯合状态存在的可能性较小,这适合用于评估遗传清除的效率。在此研究中,发现所有大熊猫种群的ROH内LoF和dnsSNP的频率显著低于ROH外(图5C),这表明许多隐性强效有害突变仍然以杂合状态存在于非ROH区域,并未有效地从熊猫种群中清除。否则,ROH与非ROH区域之间的有害突变频率将无差异,因为严重的有害突变已经从种群中清除,其余的突变应该不那么有害,或者至少不是致命的;因此,它们的频率不会受到反复近亲繁殖和纯合性的影响。
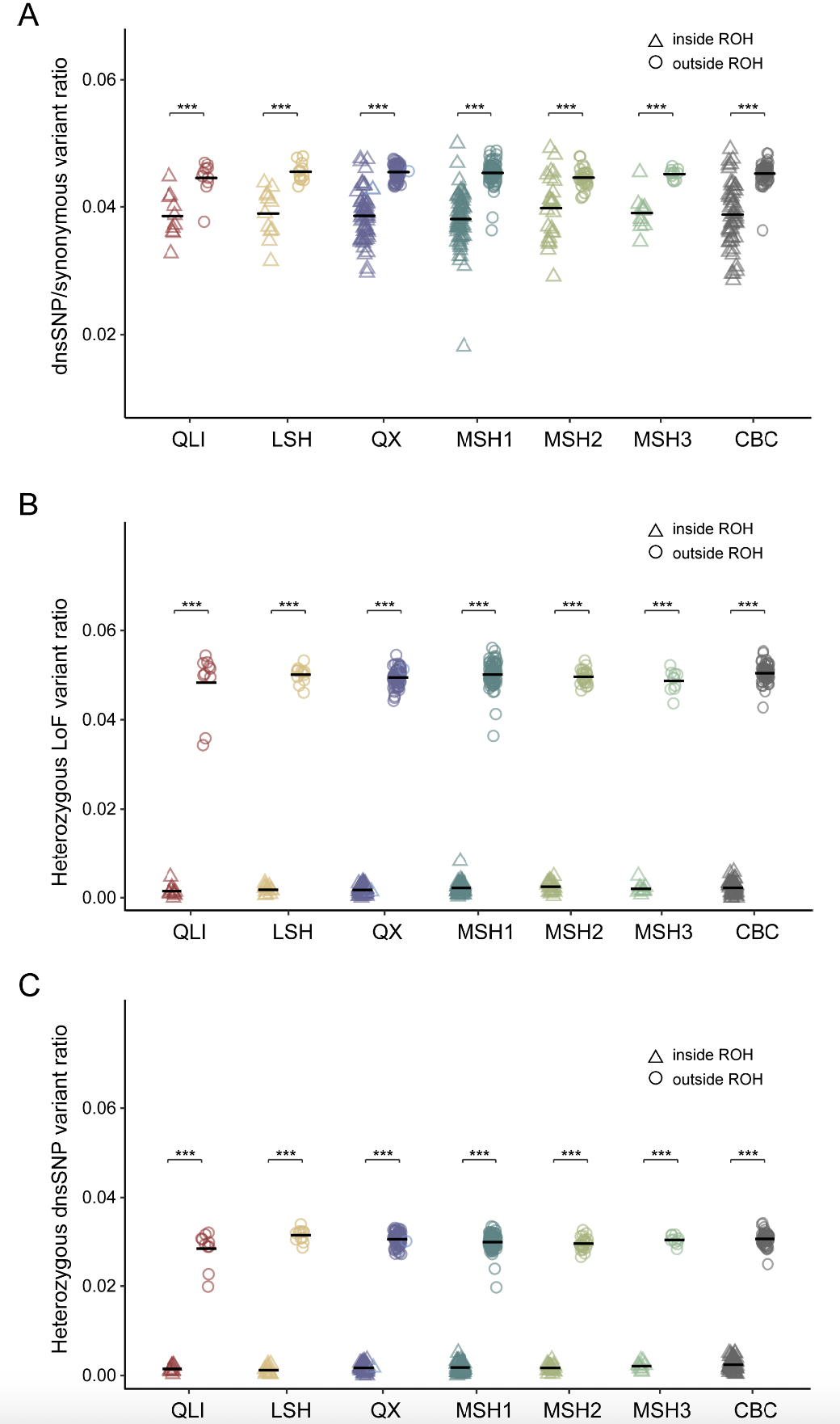
仅针对杂合有害突变的分析得出了相同的结果。大熊猫中的这种现象可能是由于低水平的近亲繁殖,不能有效揭示此类突变。然而,秦岭(QLI)和凉山(LSH)种群中ROH与非ROH区域的差异小于MSH和QX种群。这表明由于更高的近亲繁殖水平,QLI和LSH种群中的严重有害突变清除效率高于MSH和QX种群,这一点也通过 分析得到证实。
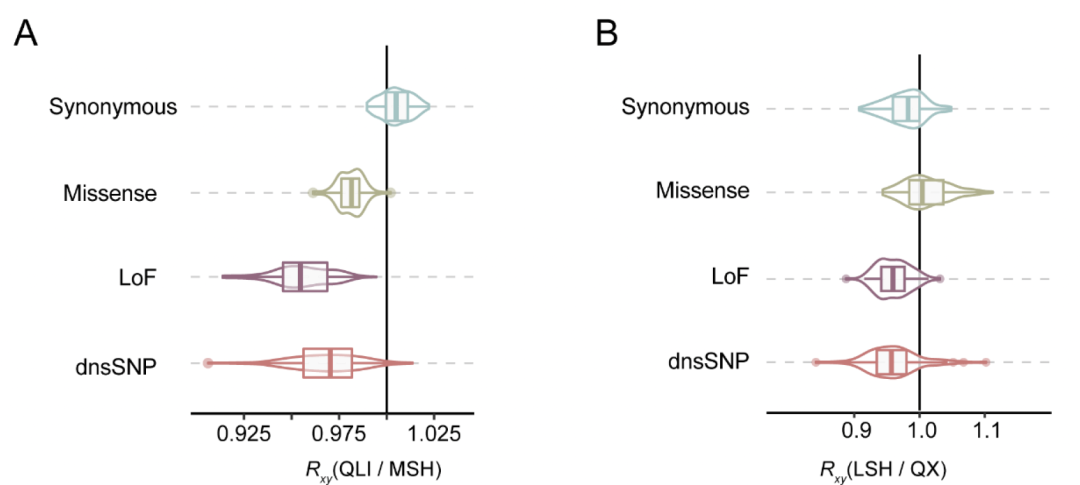
GERP分析表明,高GERP分数的大熊猫种群之间的相对突变负荷差异与低GERP分数的计算结果相似,这表明对于最有害的等位基因,遗传清除的效率并未显著增加。最后仍然发现,有害的等位基因在所有大熊猫种群中的频率低于中性等位基因,这表明纯化选择一直在发挥作用,尽管清除可能并不强烈。同样在圈养大熊猫中没有检测到明显的遗传清除,这也是意料之中的,因为圈养大熊猫的繁殖管理得到了改善(图5C)。
圈养大熊猫释放到QLI和LSH种群的模拟
为了预测引入来自不同遗传谱系(不同山脉)的圈养大熊猫后,接收的野生种群在几代内可能发生的有害突变累积,针对两个最小种群(QLI和LSH)在三种情景下进行了时间前向模拟:1)圈养大熊猫来自与接收的野生种群相同的山脉;2)杂交圈养大熊猫有一半来自接收的野生种群;3)圈养大熊猫来自不同于接收野生种群的山脉(图5D和E)。
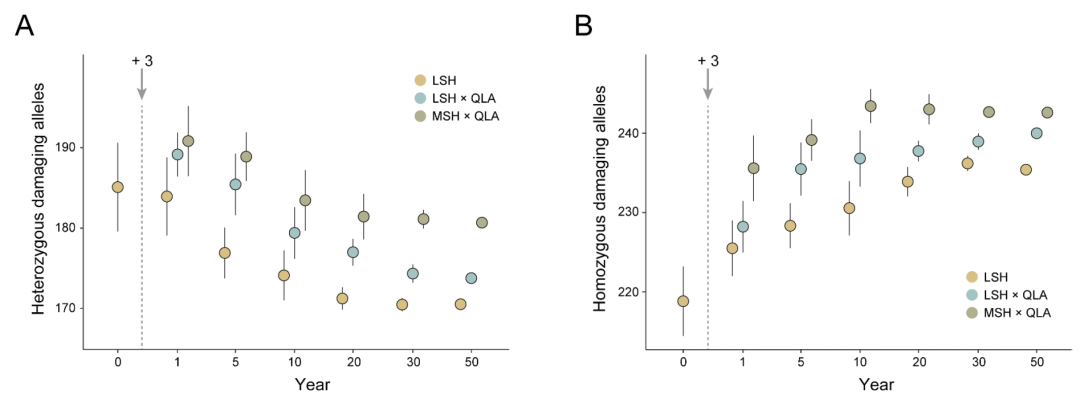
结果表明,与其他两种情景相比,引入来自与接收种群相同山脉的圈养个体似乎是最好的策略。在这三种情景下的种群恢复阶段,观察到QLI和LSH种群中杂合有害突变的减少趋势和纯合有害突变的增加趋势。有趣的是,发现杂合有害突变的数量与引入的圈养大熊猫种群(CBC种群)与接收的野生种群之间的遗传关系呈正相关,引入来自与接收种群相同山脉的圈养个体将在种群恢复期间引入最低水平的遗传负荷。
重要性
大熊猫正面临加速的栖息地碎片化,其在中国四川、陕西和甘肃省的六个山脉中形成了几个相对孤立的亚种群。然而,最近碎片化的栖息地是否导致了种群分化,以及野生大熊猫面临的风险是什么,仍然在很大程度上未知,这些知识对于未来的保护工作非常重要。在此研究中,作者进行了涵盖所有当前种群的大规模种群基因组研究,揭示了遗传结构,并在进化历史背景下提供了种群动态的图景。对遗传背景的鉴定以及对近亲繁殖和有害等位基因积累风险的评估,为大熊猫的未来保护提供了重要的知识。
文章来源:MEPGT公众号



 京公网安备11010102006642号
京公网安备11010102006642号